Название работы: Грибы. Слизевики. Особенности строения и деления на таксоны
Предметная область: Биология и генетика
Описание: Среди них есть как свободноживущие в почве на разлагающихся растительных остатках и тому подобных субстратах так и паразиты водорослей водных грибов и высших водных и наземных растений. Черты строения гифохитриомикотовых и лабирунтуломикотовых грибов.
Дата добавления: 2015-03-17
Размер файла: 2.09 MB
Работу скачали: 4 чел.
- Слизевики. Особенности строения и деления на таксоны. Отдел плазмодиофоромикотовые слизевики: характеристика основных представителей.
Под общим названием слизевики объединяется свыше 450 видов бесхлорофилльных организмов, довольно разнообразных по строению и образу жизни. Среди них есть как свободноживущие в почве, на разлагающихся растительных остатках и тому подобных субстратах, так и паразиты водорослей, водных грибов и высших водных и наземных растений. Существенная часть слизевиков обнаружена и описана только в середине 20 века [1].
Вегетативное тело слизевиков представлено тремя основными типами:
1) одноядерная или многоядерная амебоидная клетка (характерна для некоторых из слизевиков, имеющих микроскопически малые размеры);
2) плазмодий – вегетативное тело в виде крупного многоядерного цитоплазменного образования, достигающего в отдельных случаях даже нескольких десятков сантиметров;
3) псевдоплазмодий – вегетативное тело, по внешнему виду напоминающее плазмодий, но представляющее собой скопление амеб, которые не теряют своей индивидуальности, однако при этом ведут себя как единое целое. В вегетативной стадии большинство слизевиков, представленных амебоидными клетками, размножается делением этих клеток. Для некоторых слизевиков с плазмодиями показано также, что в течение вегетативной жизни от плазмодиев могут отделяться небольшие участки, далее существующие в виде двужгутиковых зооспор. Для ряда слизевиков характерно образование склероциев (утолщенная твердеющая масса), которые сохраняют жизнеспособность в течение не одного десятка лет [2].
Длительное время слизевики объединялись в один отдел, включавший классы, различающиеся по организации и особенностям циклов развития своих представителей: протостелиевые ( Protosteliomycetes ), миксогастровые, или собственно слизевики ( Myxogasteromycetes ), плазмодиофоровые ( Plasmodiophoromycetes). акразиевые, или клеточные слизевики (Acrasiomycetes ). В некоторых системах слизевики объединялись в царство простейшие (Protozoa) и делились на два (Myxomycota и Plasmodiophoromycota ) или четыре отдела ( Myxomycota, Dyctiosteliomycota, Acrasiomycota и Plasmodiophoromycota) [3].
В системе, предлагаемой Костиковым И.Ю. с соавт. (2006) слизевики на основании строения митохондриального аппарата и особенностей биохимии внутриклеточных процессов в ранге отделов разнесены по разным царствам: акразиомикотовые слизевики ( Acrasiomycota ) отнесены к царству дискокристатные (Discicristates ); миксомикотовые слизевики ( Myxomycota ), которые включают и диктиостелиевые слизевики в ранге класса – к царству трубчатокристатные ( Tubulocristates ); плазмодиофоромикотовые слизевики ( Plasmodiophoromycota) отнесены к царству пластинчатокристатные ( Platicristates). Во всех современных системах плазмодиофоромикотовые слизевики имеют ранг отдела Plasmodiophoromycota [1].
Отдел плазмодиофоромикотовые слизевики (Plasmodiophoromycota ), имеющий в своем составе один класс ( Plasmodiophoromycetes ), включает внутриклеточных паразитов, в отличие от других слизевиков не образующих органов спороношений, споры развиваются внутри клеток хозяина и имеют хитиновую клеточную оболочку.
Наиболее характерный представитель класса – плазмодиофора капустная (Plasmodiophora brassicae). Она является возбудителем болезни капустных (крестоцветных), называемой «кила». Заболевание выражается в уродливых разрастаниях корней в виде наростов и вздутий, которые со временем преобразуются в неправильной формы опухоли. При этом нарушается проводимость корней, они почти не ветвятся. Нарушение нормальной работы корневой системы приводит к гибели растения (если заражение произошло на ранних этапах его развития) либо к увяданию листьев. Зараженные растения капусты не образуют кочанов, либо они недоразвиты [3].
Другой представитель этого класса, имеющий важное практическое значение, – возбудитель порошистой парши картофеля спонгоспора (Spongospora solani). Многоядерные плазмодии спонгоспоры развиваются в периферических клетках клубня и образуют затем губчатые комочки плотно спаянных спор. Пораженный клубень покрыт язвочками с коричневым порошистым содержимым, состоящим из скоплений этих комочков. Иногда развивается корневая форма в виде небольших белых желвачков, темнеющих и затем распадающихся. Кроме картофеля поражаются томаты и другие виды пасленовых [3].
- Псевдогрибы: место в ситеме органического мира. Отдел оомикотовые грибы, характеристика отдела и основных представителей. Черты строения гифохитриомикотовых и лабирунтуломикотовых грибов.
Псевдогрибы – это гетеротрофные организмы, питающиеся осмотрофным путем, для которых характерно наличие как минимум одного локомоторного жгутика, покрытого ретронемами (трехчленными мастигонемами), начальные стадии образования которых происходят между мембранами ядерной оболочки. Гетеротрофность псевдогрибов, вероятно, вторичная, поскольку их происхождение связывается с хромофитовыми водорослями, утратившими по различным причинам фотосинтетический аппарат [2].
Под общим названием «псевдогрибы» в настоящий момент объе-диняются представители трех отделов: Оомикотовые грибы ( Oomycota ),
Гифохитридиомикотовые грибы ( Hyphochytriomycota ), Лаби-ринтуломикотовые грибы, часто именуемые также сетчатые слизевики ( Labyrinthulomycota ). Выделение отделов псевдогрибов происходит на основе данных по организации жгутикового аппарата, общему плану строения вегетативного тела, способам размножения [3].
Положение псевдогрибов в системе органического мира неоднозначно трактуется разными авторами. В учебном пособии Л. В. Гарибовой и С. Н. Лекомцевой (2005) они рассматриваются в составе самотоятельного царства Chromista. В системе И.Ю. Костикова с соавт. (2006) группа отделов псевдогрибы вошла в состав подцарства Страменопилы Stramenopiles ( царство Tubulocristates ), которое наряду с ними включает большую группу отделов хромофитовых водорослей ( Raphidophyta, Chrysophyta. Eustigmatophyta, Xanthophyta, Phaeophyta, Bacillariophyta, Dictyochophyta ) с вторично симбиотическими четырехмембранными пластидами родофитного происхождения и разножгутиковыми монадными стадиями [1].
Ключевым и наиболее широко представленным среди псевдогрибов является отдел оомикотовые грибы ( Oomycota ), который включает большую группу водных грибов, обитающих на растительных остатках, трупах животных или паразитов водорослей, других водных грибов, беспозвоночных животных, амфибий, рыб. Некоторые живут в почве. Наиболее высокоразвитые из них – облигатные паразиты высших наземных растений. Зооспоры этих грибов характеризуются двумя жгутиками примерно равной длины. Один из них снабжен мастигонемами (перистый), другой – гладкий. Состав клеточной стенки также уникален: ее основу составляют целлюлоза и глюканы, хитин отсутствует. Кроме того, кристы митохондрий имеют трубчатое строение с базальной перетяжкой. Вегетативное тело варьирует от одноклеточного образования у более примитивных форм до хорошо развитого несептированного мицелия.
Бесполое размножение осуществляется зооспорами, у немногих –конидиями (их структура принципиально отличается от конидий настоящих грибов). Половой процесс оогамный, но содержимое антеридия не дифференцировано на гаметы. Оплодотворенные яйцеклетки покрываются многослойной оболочкой и превращаются в ооспоры (зиготы) [1].
В состав отдела входит один класс с несколькими порядками, из которых важнейшими являются сапролегниальные ( Saprolegniales ), пероноспоральные ( Peronosporales) и питиальные (Pythiales ), выделенные из пероноспоральных в самостоятельный порядок. Порядок сапролегниальные ( Saprolegniales ). Большинство представителей – водные сапротрофы на остатках животных или растений. Некоторые паразитируют на икре рыб и лягушек, на рыбах, беспозвоночных животных, на морских и пресноводных водорослях, на водных грибах, на корнях высших наземных растений. У некоторых видов вегетативное тело одноклеточное, микроскопических размеров, у большинства – хорошо развитый несептированной мицелий. Бесполое размножение происходит с помощью двужгутиковых зооспор, причем у многих представителей существуют две стадии зооспор различных по внешнему виду и последовательно сменяющих друг друга через стадию цисты. Такое явление называется дипланетизм. Половой процесс оогамный, причем в оогонии образуются у некоторых одна, а у большинства много яйцеклеток, на формирование которых идет все содержимое оогония, а антеридий не дифференцирован на гаметы. Оплодотворение происходит путем переливания цитоплазмы и ядра из антеридия в яйцеклетку через оплодотворяющий отрог, отходящий от антеридия и проникающий внутрь оогония. Порядок пероноспоральные ( Peronosporales ) содержит более 500
видов грибов, весьма разнообразных по образу жизни и по морфологии. В пределах порядка ясно прослеживается эволюция от водных форм к наземным и от сапротрофного образа жизни к облигатному паразитизму [3].
От сапролегниальных грибов пероноспоральные прежде всего отличаются тем, что при формировании оогония не все его содержимое идет на построение единственной яйцеклетки, часть его остается в виде так называемой периплазмы, окружающей яйцеклетку. Периплазму можно рассматривать как прогрессивный признак, так как она обеспечивает развитие ооспоры и формирование ее оболочки. Порядок включает следующие семейства: пероноспоровые ( Peronosporaceae ), альбуговые (А lbuginaceae ) и питиевые (Pythiaceae ), которые в последнее время выделены в самостоятельный порядок питиальные (Pythiales ), отличающиеся разветвленными спорангиеносцами и зооспорангиями, способными в зависимости от условий прорастать зооспорами или гифой. Очень важное практическое значение среди питиальных имеет род фитофтора ( Phytophthora). в рамках которого известно около 70 видов. Они характеризуются более развитым и толстым мицелием, симподиально ветвящимися спорангиеносцами, которые отличаются от гиф вегетативного мицелия. Зооспорангии имеют лимоновидную, яйцевидную, шаровидную форму и сосочек на вершине, у некоторых видов вскрывающийся крышечкой. Они опадают со спорангиеносцев и так распространяются. Зооспоры формируются в зооспорангиях и 12 выходят из него поодиночке, реже одетые общим пузырем. В некоторых случаях зооспорангии прорастают гифой [1] .
- Классы тафриномицеты, схизосахаромицеты и сахаромицеты. Общая характеристика. Особенности протекания жизненного цикла, представители.
Данные классы относятся к отделу аскомицетовые грибы (Ascomycota)
Отдел аскомикотовые, или сумчатые грибы ( Ascomycota ) один из обширнейших отделов грибов, включающий более 50 000 видов (включая около 16 000 видов лишайниковообразующих грибов), с широким диапазоном жизненных форм. Сюда относятся дрожжи, представленные одиночными почкующимися клетками, многочисленные микроскопические грибы и грибы с крупными плодовыми телами, достигающими нескольких сантиметров, а у некоторых видов – 10–30 см (сморчки, строчки и др.). Все это многообразие грибов связано общим происхождением и имеет ряд общих черт [4].
Подотдел тафриномикотовые ( Taphrinomycotina ) включает четыре класса, ключевой из которых – тафриномицеты ( Taphrinomycetes). Данный класс включает представителей с псевдодикарионтическим мицелием, преобладающим в жизненном цикле. Первоначально гаплоидный мицелий формируется в результате почкования аскоспор, двухъядерность его клеток возникает в результате одного из митозов, вследствие чего клетки мицелия содержат генетически однородные ядра, и только морфологически напоминают дикарион. Образованию аскоспор предшествует слияние ядер одной из клеток (кариогамия) и последующий мейоз (представители: тафрина ( Taphrina). протомицес (Protomyces )). К этому же отделу на основа-нии молекулярных и цитогенетичеких данных относится и недавно выделенный класс схизосахаромицеты ( Schizosaccharomycetes ), объединяющий делящиеся, гаплоидные дрожжи (центральный род – схизосахаромицес ( Schizosaccharomyces ) [3].
Подотдел сахаромикотовые ( Saccharomycotina ) включает один класс сахаромицеты (Saccharomycetes), объединяющий почкующиеся диплоидные и гаплоидные дрожжи. Наиболее яркие представители: сахаромицес (Saccharomyce s) и сахаромикодес ( Saccharomycodes ).
Среди аскомикотовых грибов основная часть дрожжей относится к классам схизосахаромицеты ( Schizosaccharomycetes ) и сахаромицеты ( Saccharomycetes ). Ранее эти группы входили в состав одного порядка эндомицетовые, но в настоящий момент разнесены в разные подотделы. Общим для них является образование одиночных сумок, развивающихся из зиготы, без участия аскогенных гиф и дикарионтической ядерной фазы вообще. Мицелий отсутствует [3].
Класс схизосахаромицеты ( Schizosaccharomycetes) объединяет делящиеся гаплоидные дрожжи (род схизосахаромицес – Schizosaccharomyces ), у которых половой процесс протекает в виде копуляции гаплодных соматических клеток, псевдомицей не формируется. Дрожжи класса сахаромицеты ( Saccharomycetes ) – это почкующиеся представители. Чередование ядерных фаз и способы полового размножения разнообразны. Например, у сахаромикодеса Людвига ( Saccharomycodes ludvigii ) гаплоидная ядерная фаза короткая, диплоидизация происходит в виде слияния аскоспор, часто уже в сумке (рис 1) [1].
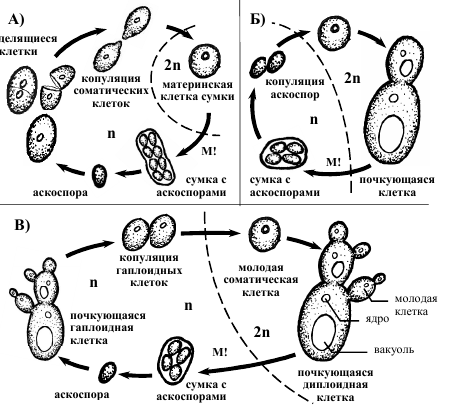
Рисунок 1 – Схема жизненных циклов сумчатых дрожжей: а) схизосахаромицес Schizosaccharomycetes ; б) сахаромикодекс Людвига Saccharomycodes ludvigii ; в) пекарские дрожжи Saccharomyces
- Классы эвроциомицеты и сордариомицеты. Общая характеристика. Порядки сордариальные и гипокреальные. Характеристика представителей.
Класс эвроциомицеты ( Eurotiomycetes ) включает грибы с плодовыми телами в виде клейстотециев, внутри которых прототуникатные сумки разбросаны беспорядочно (см. рисунок 10). Плодовые тела шаровидной формы, светлоокрашенные, тонкостенные. Аскоспоры освобождаются пассивно после разрушения оболочки плодового тела. У ряда видов преобладает анаморфная стадия, а часто сумчатая стадия (телиоморф) редуцируется вообще. Наиболее яркие представители: эвроциум ( Eurotium ) и эмерицелла ( Emericella ), анаморфные стадии которых имеют собственные названия и отнесены к формальным родам несовершенных грибов Penicillum и Aspergillus. Под названием пеницилл объединяются конидиальные стадии и других видов эвроциевых ( Eupenicillum, Talaromyces ), которые отличаются кистевидным строением конидиеносца. Его строение разнообразно, но в наиболее развитом виде конидиеносец состоит из ветвей, на которых расположены метулы с мутовками фиалид. На фиалидах образуются цепочки конидиоспор [1].
У аспергилла конидиеносец одноклеточный или с поперечными перегородками. На верхушке конидиеносец вздут и образует пузырь. На поверхности вздутия расположены короткие фиалиды с цепочкой отделяемых конидий (фиалоконидий). У некоторых аспергиллов фиалиды находятся не на самом вздутии, а на специализированных клетках – профиалидах [2].
Сумчатые стадии у ряда представителей формальных родов Penicillum, Aspergillus. а также Acremonium отсутствует или еще не обна ружены, поэтому большое количество грибов со сходным бесполым спороношением относится к формальному отделу дейтеромикотовые грибы ( Deuteromycota ). При обнаружении у них сумчатой стадии или установления родства цитогенетическими методами виды на полных правах относятся к эвроциомицетам. Зачастую виды даже после этого сохраняют тривиальное название по конидиальной стадии, поскольку половое спороношение образуется редко, а без него определение вида требует инструментального цитогенетического анализа [4].
Класс сордариомицеты ( Sordariomycetes ). Представители имеют плодовые тела в виде микроскопических перитециев, расположенных на мицелии, как у сордарии и цератоцистиса, или в стромах. Сумки унитуникатные, иноперкулятные. Есть конидиальное спороношение. Ключевые порядки: сордариальные (Sordariales), ксилариальные ( Xylariales) и гипокреальные (Hypocreales ). Ранее выделяемый порядок спорыньевые, или клавицепсовые ( Clavicipitale s) в представленной систематике вошел в состав гипокреальных. Половые органы некоторых грибов данного класса настолько упрощены, что зачастую их половой процесс определяется как переходная форма между гаметангиогамией и соматогамией (например, у некоторых видов рода Sordaria).
Порядок гипокреальные (Hypocreales ) объединяет виды, которые образуют перитеции в хорошо развитых стромах, либо на мицелиальных сплетениях. Стромы обычно мясистые, яркоокрашенные, распростертые по субстрату, подушковидные или головчатые. Стромы развиваются на субстрате – обычно на пораженных органах растенияхозяина или из склероциев (виды спорыньи – Claviceps ), или из мумифицированных, пронизанных гифами гриба тканей хозяина – псевдосклероциев (виды рода кордицепс – Cordyceps ). Лишь у немногих представителей этого порядка строма отсутствует. Перитеции имеют типичное строение, погружены в стромы, так что на поверхность выступают только носики перитециев. К моменту созревания аскоспор в верхней утолщенной части сумки образуется пора, через которую они выходят наружу. Споры расположены в сумке параллельным пучком и выбрасываются поочередно. Большинство представителей порядка – паразиты на цветковых растениях, грибах и членистоногих. Лишь очень немногие обитают как сапротрофы на почве или на древесине.
Ключевые представители порядка: нектрия ( Nectria ), спорынья ( Claviceps), кордицепс (Cordyceps), эпихлое (Epichloe), гибберелла (Gibberella). Наиболее распространенный из них – спорынья пурпурная ( Claviceps purpurea ), развивающаяся на многочисленных видах злаков, как культурных, так и дикорастущих (рис 7) [3].
- Класс пецицомицеты. Порядки пецицальные и туберальные. Представители. Строение плодовых тел. Класс дотидеомицеты (локулоаскомицеты) Особенности строения псевдомицетов. Строение сумок и сумкоспор. Основные представители.
Класс пецицомицеты (Pezizomycetes ) включает представителей с макроскопическими апотециями, сумки унитуникатные, оперкулятные. Стромы и конидиальное спороношение не характерны. Ключевые порядки: пецицальные (Pezizales) и туберальные (Tuberales) [1].
Порядок пецицальные (Pezizales) характеризуется сумками, открывающимися на вершине крышечкой и плодовыми телами в виде типичных апотециев (рисунок 20) от очень мелких, не превышающих 1 мм в диаметре, до крупных, размером до 10 см. Реже образуются апотеции со стерильной ножкой: гельвеллоидные (гимений на верхней поверхности лопастной шляпки) и моршеллоидные (гимений на верхней поверхности складчатой шляпки). Такие апотеции достигают в высоту 10–12 см и более.
Апотеции имеют мясистую, реже студенистую или кожистую консистенцию. Их окраска разнообразна: от яркой, оранжевой или красной у одних представителей до коричневой или черной у других. В гимении пецицевых всегда присутствуют парафизы. Обычно по длине они равны сумкам, но у некоторых выступают за пределы гимения. Концы парафиз часто расширены и окрашены, у некоторых представителей они ветвятся. Сумки ряда пецицевых при созревании удлиняются, и выступают над гимением. Пецицевые, как правило, сапротрофы, лишь немногие могут па разитировать на растениях [1].
Для рода пецица (Peziza ) характерны типичные блюдцевидные или чашевидные апотеции размером 1–5 см, бурого или коричневого цвета, снаружи гладкие или мучнистые. Представители этого рода встречаются преимущественно в лесах на влажной почве ( Р. badia, P. pustulata и др.). Немногие из них лигнофилы, как, например, пецица фиолетово-черная (Р. violaceo-nigra ), развивающаяся на гнилой древесине и пнях лиственных деревьев. В этом роде есть также карбофилы и копрофилы. Весной и летом в лесах, особенно на старых кострищах, нередко можно встретить крупные фиолетово-коричневые апотеции пецицы фиолетовой (Р. violacea ). Другой распространенный вид этого рода пецица коричневая (Р. badia ) встречается с лета до осени на влажной почве в хвойных лесах, вдоль дорог, на опушках. Ее крупные каштаново-коричневые апотеции обычно собраны большими группами [2].
У видов рода сморчок (Morchella ) апотеции крупные, не менее 6–10 см высотой, мясистые, четко разграничены на ножку и шляпку. Шляпка правильных очертаний: яйцевидная, коническая, с сетью складок, как продольных, так и поперечных, часто косых. Складки образуют ячейки, выстланные гимением. Разделяющие их ребра остаются стерильными. Края шляпки срастаются с ножкой, внутри она полая.
В противоположность большинству крупных пецицевых у сморчков не наблюдается «взрыва», когда сразу много сумок в гимении, моршеллоидные апотеции с участками гимениев гельвеллоидный апотеций выстреливают споры. Выбрасывание аскоспор у сморчков происходит постепенно и регулируется интенсивностью солнечной радиации. Наиболее распространены два вида сморчков – сморчок съедобный (М. esculenta ) и сморчок конический (М. conica).
Для рода строчок (Gyromitra ) характерны крупные апотеции не правильных очертаний. Шляпка неправильно-яйцевидная или бес-форменная, с неупорядоченной складчатостью, бурая или темно-бурая, реже более светлая. Ножка толстая, неправильной формы, часто бороздчатая, белая или светлая. Виды этого рода – сапротрофы на почве, обильно развивающиеся весной. Наиболее распространенный вид этого рода – строчок обыкновенный, или съедобный (G. esculenta ), часто в массе развивающийся весной на почве в лесах, преимущественно сосновых. Иногда встречается до осени. Строчок обыкновенный считают условно съедобным грибом [1].
Порядок трюфелевые, или туберальные (Tuberales). К порядку 65
относится около 100 видов, образующих подземные плодовые тела, в зрелом состоянии вторично замкнутые. Плодовые тела имеют клубневидную форму, их размеры колеблются от 1 до 10 см, реже более крупные. Перидий плодового тела плотный, кожистый, его поверхность гладкая или покрыта бородавками разных размеров. Внутренняя ткань плодового тела на разрезе имеет мраморный рисунок из чередующихся светлых и темных полос. Их называют соответственно внутренними и наружными венами. Сумки трюфелевых располагаются в плодовых телах гимениальным слоем или гнездообразно. Освобождение аскоспор у этой группы всегда пассивное, после разрушения плодового тела или поедания его животным. Изучение
развития плодовых тел трюфелевых показало, что они закладываются в виде блюдцевидных образований, на вогнутой складчатой поверхности которых формируются зачатки гимения. В дальнейшем такое плодовое тело, находясь в почве, не может расти вширь, на поверхности его гимения развиваются многочисленные складки, а позднее оно замыкается. Складки превращаются во внутренние вены, а щели между ними – в наружные вены, которые заполняются рыхлой тканью из парафиз. У основания парафиз позднее образуются сумки. Трюфелевые – обязательные микоризообразователи. Наиболее ценный представитель этого порядка – черный французский трюфель (Tuber melanosporum ) формирует микоризу с дубом, буком и грабом [4].
В некоторых районах нашей страны встречается другой вид этого рода – летний трюфель (Т. aestivum), образующий микоризу с теми же деревьями. Его плодовые тела довольно крупного размера, покрыты крупными бородавками черновато-бурого цвета. Мякоть плодового тела желтовато-белая, с многочисленными буроватыми и беловатыми плотными жилками. Гриб съедобен, но по качеству значительно уступает черному французскому трюфелю [1].
Подкласс локулоаскомицеты – Loculoascomycetidae
Главные признаки подкласса – отсутствие настоящих парафиз и собственной стенки вокруг полостей (локул), закладывающихся обычно в стромах в виде камер-пиреноидов. Подкласс локулоаскомицеты делится на 5 порядков. Основные из них следующие.
Порядок мириангиевые – Myriangiales. Сумки возникают в одиночных локулах, беспорядочно расположенных в строме. Небольшая группа, объединяющая тропические и субтропические виды, преимущественно пара-зитирующие на растениях, насекомых и грибах.
Порядок дотидейные – Dothideales. Сумки образуются в локулах, расположенных в выступающих из ткани субстрата или полностью поверхностных стромах, окрашенных в темный цвет. Многие представители
порядка обитают сапротрофно на растительных остатках: отмерших стеблях и листьях, ветвях и т.д. Среди них есть и паразиты растений, например виды рода микросферелла –Mycosphaerella.
Порядок плеоспоровые – Pleosporales. Аскостромы развиваются по типу «плеоспора» и содержат одну или несколько локул. К порядку принадлежат многие широко распространенные сапрофиты на растительных остатках, например виды рода плеоспора – Pleospora, развивающиеся на отмерших частях травянистых растений, и виды рода кукурбитария – Cucurbitaria, образующие аскостромы на отмерших ветвях различных деревьев и кустарников [1].
- Отдел базидиомикотовые грибы. Общая характеристика. Мицелий первичный и вторичный. Образование базидии. Типы базидий. Принципы деления на классы.
Отдел базидиомикотовые грибы ( Basidiomycota) насчитывает более 30 000 видов, которые характеризуются наличием вегетативного тела в виде многоклеточного мицелия (гаплоидного первичного и дикарионтичного вторичного) и полового спороношения в виде базидиоспор. Монадные стадии отсутствуют. Основные компоненты многослойной клеточной стенки – хитин и глюканы. Септы между клетками простые (с простой порой) или долипоровые (с бочковидным расширением вокруг поры) [2].
У базидиальных грибов встречается вегетативное, собственно бесполое и половое размножение. Вегетативное размножение осуществляется с помощью неспециализированных участков мицелия либо особых спор образующихся на дикарионтическом мицелии (эцидиоспоры, оидиоспоры, уредоспоры и т. д.). Собственно бесполое размножение встречается достаточно редко и представлено конидиальным спороношением.
Половых органов у базидиальных грибов не образуется. Половой процесс, в основном, осуществляется путем слияния двух вегетативных клеток гаплоидного мицелия, вырастающего из базидиоспор (соматогамия). Встречается половой процесс в виде слияния базидиоспор или продуктов их почкования ( Tilletia). слияния гаплоидных клеток базидии без образования базидиоспор ( Ustillago). половой процесс в виде сперматизации (Puccinia ).
У гомоталличных видов могут сливаться гифы одного и того же мицелия. У гетероталличных, к которым относится большинство базидиальных грибов, сливаются клетки гиф, берущих начало от спор противоположных половых знаков: «+» и «–». При этом происходит слияние цитоплазмы, а ядра объединяются в пары – дикарионы, которые затем синхронно делятся. Такой дикариотичный мицелий может существовать длительное время [4].
Образование базидий - При синхронном делении пары ядер дикариона одновременно начинает формироваться пряжка в виде небольшого бокового выроста, причем оси деления ядер лежат на одном уровне и вдоль клетки. В результате деления в клетке образуются четыре ядра.
Сама клетка также делится перегородкой, разделяющей одну пару ядер: одно ядро оказывается в верхней (апикальной) клетке, другое – в нижней клетке (клетке-ножке). Из оставшейся пары ядер одно остается в верхней клетке, а второе попадает в боковой вырост – пряжку, еще не отделенную перегородкой.
В дальнейшем пряжка отделяется перегородкой и от апикальной клетки, а вершина ее, загибаясь книзу, прирастает к клетке-ножке. Между ними образуется отверстие, через которое ядро из пряжки переходит в клетку-ножку, восстанавливая ее двухъядерность. Благодаря этому клетка-ножка способна к дальнейшему функционированию: она может развивать дикариотичный мицелий и базидии. В верхней клетке также остается одна пара несестринских ядер. В дальнейшем она способна образовать базидию, при этом ядра сливаются, и диплоидное ядро редукционно делится. Сама клетка обычно вытягивается, на ее вершине формируются выросты – стеригмы, которые, вздуваясь на конце, развиваются в базидиоспоры. В каждую базидиоспору через узкий просвет стеригмы проникает по одному ядру. При созревании базидиоспоры отбрасываются со стеригмы на небольшое расстояние [1].
По своему развитию базидия напоминает сумку и гомологична ей. Схема нарастания дикариотичного мицелия и развития базидии происходит с образованием пряжки, которая также гомологична структуре аскомикотовых грибов – крючку.
холобазидии (одноклеточные) и фрагмобазидии (разделенные септами – продольными или поперечными). Ранее выделяемый тип гетеробазидия в настоящий момент рассматривается как одна из форм фрагмобазидии.
Холобазидии – одноклеточные базидии, чаще цилиндрической, булавовидной, овальной или неправильной формы. При этом если ба-зидиоспоры располагаются на одном уровне, базидия является акроспоровой (характерна для грибов с активным отбрасыванием базидиоспор, а если на разных уровнях, беспорядочно –плевроспоровой (у грибов с пассивным распространением базидиоспор.
Фрагмобазидия – базидия, разделенная поперечными перегородками на две или четыре клетки, на которых формируются базидиоспоры. Их структура разнообразна и специфична для таксонов [1].
Систематика класса базидиомицеты является морфологической и построена на основании фенотипических признаков и цитологических признаков, основные их них: строение и способ образования базидии, тип и консистенция плодового тела, строение гимения, тип гименофора, наличие перфораций в парентосоме, наличие или отсутствие пряжки на дикарионтичном мицелии, орнаментация оболочки споры и ее химический состав [3].
Отдел Basidiomycota принято делить на три класса:
1) класс базидиомицеты (Basidiomycetes );
2) класс телиомицеты ( Teliomycetes). или ржавчинные грибы ( Uredinomycetes );
3) класс устомицеты ( Ustomycetes ) или головневые грибы (Ustillaginomycetes ).
В зависимости от сочетания перечисленных признаков выделяется до 14 порядков, которые объединяются в четыре несистематические морфологические группы:
– афилофороидные гименомицеты, включают порядки полипоральные ( Polyporales ), кантареллальные, или лисичковые ( Cantarellales ), телефоральные ( Telephorales ), гименохетальные (Hymenochaetales ) и фистулинальные ( Fistulinales );
– агарикоидные гименомицеты объединяют болетальные ( Boletales ), агарикальные ( Agaricale s) и руссулальные, или сыроежковые (Russulales );
– гастеромицеты: дождевиковые ( Lycoperdales ), ложнодождеви-
ковые ( Sclerodermatales ), нидуллариальные, или гнездовковые ( Nidulariales ) и фаллальные, или веселковые ( Phallales );
– фрагмобазидиомицеты со студенистыми плодовыми телами (ранее выделялись в таксон под названием гетеробазидиомицеты), которые объединяют порядки аурикулальные ( Auricullales) и тремеллальные, или дрожалковые ( Tremellales ).
- Общая характеристика лишайников. Типы строения таллома. Основные систематические группы.
Лишайники —это своеобразная группа живых организмов, тело (слоевище) которых образовано двумя организмами—грибом (микобионт) и водорослью или цианобактерией (фикобионт), находящимися в симбиозе. В составе лишайников обнаружено около 20 тыс. видов грибов и около 26 родов фототрофных организмов. Наиболее часто встречаются зеленые водоросли родов требуксия, трентеполия и цианобактерия носток, являющиеся авто-трофными компонентами примерно у 90% всех видов лишайников [5].
Симбиотические (мутуалистические) взаимоотношения между компонентами лишайников сводятся к тому, что фикобионт снабжает гриб созданными им в процессе фотосинтеза органическими веществами, а получает от него воду с растворенными минеральными солями. Кроме того, гриб защищает фикобионт от высыхания. Такая комплексная природа лишайников позволяет им получать питание из воздуха, атмосферных осадков, влага росы и туманов, частиц пыли, оседающей на слоевище, из почвы. Поэтому лишайники обладают уникальной способностью существовать в крайне неблагоприятных условиях, часто совершенно непригодных для других организмов, — на голых скалах и камнях, крышах домов, заборах, коре деревьев и др [5].
Микобионт отличается специфичностью, т. е. входит в состав только одного вида лишайника.
Слоевище лишайников обычно серого, светлого или темно-бурого цвета. По внешнему виду талломы лишайников делятся на накипные, листоватые и кустистые.
Наиболее распространены накипные, или корковые, лишайники (около 80%), имеющие таллом в виде тонкой корочки, прочно срастающейся с субстратом и не отделимой от него. Более высокоорганизованные листоватые лишайники имеют вид чешуек или пластинок, прикрепляющихся к субстрату пучками гиф, называемых ризинами. Они разрастаются на камнях и коре деревьев. Так, например, на стволах и ветках осины часто встречается лишайник золотистого цвета—ксантория. Кустистые лишайники представляют собой кустики, образованные тонкими ветвящимися нитями или стволиками, прикрепленными к субстрату лишь основанием.
По анатомическому строению лишайники делят на гомео- и гетеромерные. У гомеомерных лишайников слоевище представляет собой рыхлое сплетение гиф гриба, среди которых более или менее равномерно располагаются клетки или нити фикобионта.
Гетеромерное строение характеризуется наличием в слоевище дифференцированных слоев, каждый из которых выполняет определенную функцию: верхняя и нижняя кора — защитную, фотосинтезирующий слой участвует в процессе фотосинтеза и накапливает продукты ассимиляции, а сердцевина — в прикреплении таллома к субстрату и обеспечении аэрации фикобионта. Такой морфологический тип лишайника представляет собой наиболее высокоорганизованную форму слоевища и характерен для большинства листоватых и кустистых лишайников.
Размножение. Размножаются лишайники главным образом вегетативным путем — частями слоевища, а также особыми специализированными образованиями — соредиями и изидиями [5].
Основные систематические группы:
Класс Фиколихнес – Phycolichenes. Микобионты – грибы с неклеточным мицелием, фикобионты – синезеленые водоросли из рода Nostoc.
Класс Сумчатые – Ascolichenes. Микобионты – сумчатые грибы. Плодоношение – перитеции или апотеции с разным типом онтогенеза.
Подкласс Пиренокарповые – Pyrenocarpeae. Подкласс объединяет один порядок – перенокарповые –Pyrenocarpales. Плодовые тела – перитеции. Род Dermatocarpon объединяет лишайники с чешучатыми или листоватыми слоевищами.
Подкласс Гимнокарповые – Cymnocarpeae. Плодовые тела – апотеции, реже гастеротенного типа.
Серия порошковатоплодные – Coniocarpiidae. Порядок калициевые – Caliciales. Апотеции на слоевищах располагаются на ножках разной высоты или разветвленных прямостоящих кустиках. Слоевище накипные, листоватые, кустистые. Виды рода калициум –Calicium и кониоцибе – Coniocybe.
Порядок артониевые – Arthoniales. Порядок объединяет формы с апотециями и гастеротециями, особенность которых – отсутствие слоевищного и собственного края.
Порядок графидовые – Graphidales. Плодовые тела в виде гастеротециев, имеющих форму простых или разветвленных линий. Слоевище накипное. Графис написанный – Graphis scripta.
Серия круглоплодные – Cyclocarpiidae. Роды коллема ( Collemа) и лептогиум (Leptogium), Род лобария (Lobaria), Род лецидея (Lecidea), Род кладония (Cladonia), кладина (Cladina), стереокаулон (Stereokaulon), Род гипогимния (Hypogymnia) и пармелия (Parmelia), Род уснея (Usnea).
Класс Базидиальные лишайники – Basidiolichenes. Класс включает около 20 видов лишайников, у которых микобионты – грибы из порядков Aphyllophorales и Agaricales.
Класс Дейтеролихенес – Deuterolichenes (Lichenes imperfecti). Класс включает лишайники со стерильным слоевищем, размножающиеся соредиями. Род лепрария – Lepraria [3,5].
- Жизненный цикл сапролегнии ( Saprolegniа)
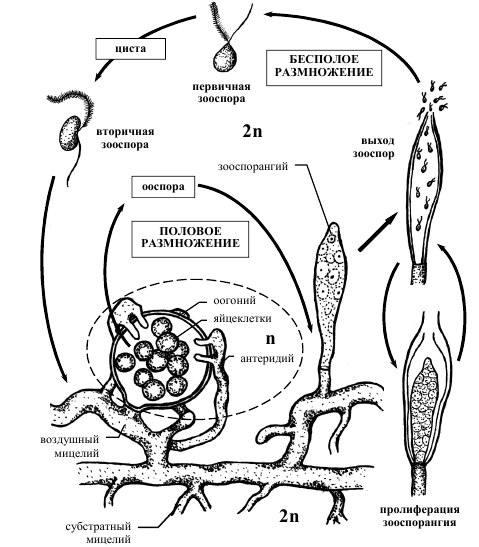
Рисунок 2 – Схема жизненного цикла представителей рода сапролегния Saprolegniа
У видов рода Saprolegniа вегетативное тело представлено хорошо 6+развитым несептированным (неклеточным) мицелием, состоящим из субстратного мицелия (более тонкие ризоидальные гифы), пронизывающего субстраты и воздушного мицелия (более толстые вегетативные гифы), несущего органы бесполого и полового размножения (рисунок 2).
Бесполое размножение осуществляется зооспорами, которые формируются в зооспорангиях булавовидной формы. Сами зооспорангии образуются из утолщенных концов вегетативных гиф, цитоплазма которых сгущается и отделяется от мицелия перегородкой. После выхода из них через верхушечное отверстие зооспор в опустевший зооспорангий может врастать новый зооспорангий от той же гифы (пролиферация), этот процесс может повторяться несколько раз Зооспоры, вышедшие из зооспорангия, являются первичными, имеют грушевидную форму и несут два жгутика на переднем конце [1].
Поплавав некоторое время (около получаса), они останавливаются, одеваются оболочкой и переходят в состояние покоя (циста), а затем
прорастают, выпуская почковидную вторичную зооспору со жгутиками, прикрепленными сбоку. Эти зооспоры имеют обычно более длительный период плавания. Найдя подходящий субстрат, они садятся на него, втягивают жгутики, одеваются оболочкой и прорастают в мицелий.
При половом процессе на мицелии развиваются оогонии и анте-ридии. Оогонии представляют собой шаровидное вздутие на короткой боковой ветви, отделенное от нее перегородкой, где развивается несколько яйцеклеток, на построение которых идет весь протопласт оогония. Антеридии представляют собой небольшие конечные многоядерные участки так называемых антеридиальных ветвей мицелия, также отделенные перегородками. У одних сапролегниевых грибов они развиваются на том же мицелии, что и оогонии (гомоталличные виды), a у других – на другом (гетероталличные виды). Антеридиальные ветви подрастают к оогонию. Антеридий плотно прикладывается к его оболочке и пускает оп-лодотворяющие отроги (один или несколько) внутрь оогония через поры, заметные в виде маленьких кружочков в оболочке оогония. Ядро и цитоплазма антеридия попадают в яйцеклетку. Один антеридий может оплодотворить одну или несколько яйцеклеток. После оплодотворения развивается ооспора, одевающаяся толстой оболочкой. Таллом сапролегниевых грибов – диплоидное образование, а число хромосом редуцируется при формировании гамет. Ооспора прорастает зооспорангием или короткой гифой с зооспорангием.
- Жизненный цикл фитофторы инфекционной ( Phytophthora infestans)
Мицелий гриба распространяется преимущественно по межклетникам листа, а в клетки внедряет гаустории. Пораженные участки быстро отмирают, и на листьях появляются бурые пятна отмершей ткани. По краю такого пятна с нижней стороны листа бывает хорошо заметен, особенно во влажную погоду, беловатый пушок. Он представляет собой скопление спорангиеносцев (конидиеносцев), высовывающихся из устьиц целыми пучками. Они симподиально ветвятся и несут лимоновидные зооспорангии, которые всегда отваливаются целиком и переносятся на новый лист или с каплями дождя попадают через почву на клубни. В капле воды они прорастают зооспорами, которые после некоторого периода движения округляются, а затем развивают гифы, проникающие или внутрь листа (через устьица или непосредственно через эпидермис), или в клубень. В сухую погоду спорангии могут прорастать прямо в гифы, т. е. фактически по способу прорастания превращаются в конидии. Образование спорангиеносцев, прорастание зооспорангиев (или конидий, как их чаще называют) и заражение растений происходят главным образом ночью или под утро. В дождливую погоду или при обильных росах и туманах ботва гибнет в течение 7–10 дней на десятках и сотнях гектаров. Массовое заражение клубней наблюдается в основном при уборке картофеля. На клубнях появляются пятна свинцово-серого цвета. При хранении недостаточно просушенных клубней в условиях повышенной влажности и температуры они начинают гнить. Присутствие паразита стимулирует амилазу клубня, и крахмал переходит в сахар. На зараженных клубнях обильно развиваются бактерии, и гниль из сухой превращается в мокрую (рис 3) [1].
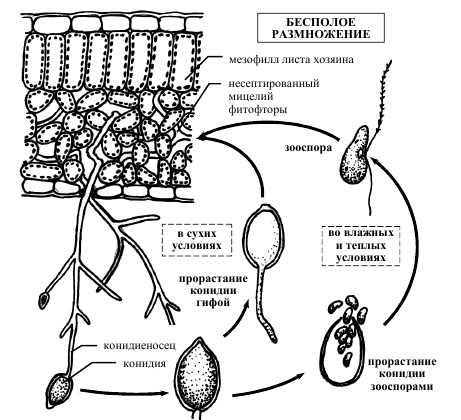
Рисунок 3 – Схема бесполого размножения фитофторы Phytophthora infestans.
Половой процесс наблюдается редко, потому что гриб гетероталличен, и для его протекания необходимо присутствие штаммов обоих типов спаривания [1].
Меры борьбы с фитофторозом:
Для повышения устойчивости растений к болезни увеличивают дозы калийных удобрений.
При обнаружении первых признаков фитофторы на картофеле, необходимо провести профилактическое опрыскивание одним из препаратов: арцерида, каптана (0,5%), поликарбацина (0,4%), полихома (0,4%), ридомила (0,2%), хлорокиси меди (0,4%) или цинеба (0,4%).
Рассаду томата и перца начинают опрыскивать с фазы 2-3 листьев агатом-25К с интервалом 25 дней [5].
При сильном поражении томата фитофторозом проводят немедленный сбор плодов с их дальнейшим дозариванием.
Растительные остатки необходимо удалять с грядки.
Важно соблюдать правила севооборота при выращивании томата и картофеля. Недопустимо размещение этих культур в огороде рядом.
Можно подобрать сорта картофеля и томата, устойчивые к фитофторозу.
Некоторые огородники для предотвращения фитофтороза помидоров снижают влажность воздуха в теплице и повышают там температуру до +30С (закрывают теплицу в солнечный день на 1-2 часа "прогревая" в ней воздух).
Важно подвергать семена томатов предварительной обработке перед посевом. Можно выдержать их 20 минут в растворе марганцовки (1 г на 100 мл воды), потом промыть и подсушить.
Кусты томата периодически опрыскивают настоем чеснока с марганцовкой (1,5 стакана измельченного чеснока + 1,5 г марганцовки на 10 л воды).
Опытные огородники советуют опрыскивать томаты и разведенными дрожжами (100 г на 10 л воды), и настоями сосновой хвои, и раствором борной кислоты (1 чайная ложка на 10 л воды), а также раствором йода (40 капель) + 30 г хлористого калия + 10 л воды.
Можно бороться с фитофторой настоем гриба-трутовика - 100 г гриба измельчить и залить кипящей водой, накрыть крышкой и настоять. Процедить и опрыскать настоем томаты по листьям. Обработку проводят в конце июня - начале июля.
Ранние сорта картофеля успевают созреть до развития фитофторы. Устойчивые к болезни сорта: Луговская, Гранола.
Эффективны против фитофторы экологически чистые биофунгициды, например, "Фитоспорин", которым поливают растения при первых признаках болезни.
Некоторые опытные огородники проводят первое профилактическое опрыскивание растений от фитофтороза в начале июля, используя контактный фунгицид. Вторую обработку проводят через 10 дней препаратом татту. Третью - через 7 дней после второй препаратом квадрис + любое микроудобрение. Четвертую обработку проводят через 10 дней опять препаратом татту. Во время сбора плодов химические препараты не используют. Можно каждые 10 дней опрыскивать растения настоем древесной золы (полведра золы залить доверху водой и настаивать 2 дня, потом довести объем до 30 литров и добавить 100 г хозяйственного мыла).
- Жизненный цикл Плазмодиофоры капустной (Plasmodiophora brassicae ).
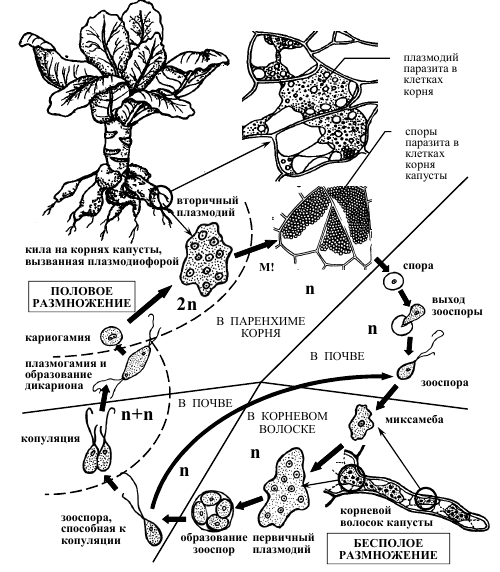
Рисунок 4 – Схема жизненного цикла плазмодиофоры капустной Plasmodiophora brassicae.
Споры плазмодиофоры сохраняются в почве годами (до 7–10 лет). Их распространению способствуют животные (дождевые черви, насекомые), токи воды (дождь, полив), деятельность человека (например, на орудиях обработки почвы). При подходящих условиях влажности и температуры, а также при стимулирующем действии корневых выделений растений-хозяев споры прорастают зооспорами с двумя жгутиками (расположены на переднем конце, гладкие, неравной длины) [2].
Зооспоры проникают в корневые волоски хозяина, где превращаются в миксамебы. Миксамебы способны сливаться без слияния ядер (плазмогамия), что приводит в формированию первичных гаплоидных плазмодиев, количество ядер в которых несколько увеличивается за счет митотического деления ядер. Из первичного плазмодия вновь могут образовываться зооспоры, которые способны выходить в почву и заражать новые корневые волоски. При определенных условиях зооспоры попарно копулируют с образованием двуядерной клетки, последняя проникает в клетки более глубоких тканей корня, где преобразуется во вторичный плазмодий, что сопровождается кариогамией и переходом в диплоидное состояние (рис 4).
Вторичный плазмодий более жизнеспособен, его протопласт разрастается, часто отчленяет новые плазмодии, проникающие в соседние клетки. Под его действием к пораженным участкам обильно притекают пластические вещества хозяина, клетки которого усиленно делятся и гипертрофируются, в результате чего получаются крупные опухоли. Через некоторое время происходит редукционное деление ядер, и плазмодий распадается на многочисленные (до 25 000), покрытые плотной оболочкой споры. Споры высвобождаются после сгнивания корней. Таким образом, часть жизненного цикла плазмодиофоры проходит в корневых волосках, часть – в почве, часть – в клетках более глубоких тканей корня [1].
4. Жизненный цикл ольпидиума капустного (Olpidium brassicae)
Заражение растения происходит зооспорами,которые при наличии воды прилипают к поверхности стебелька капусты, утрачивают жгутики и переливают свое содержимое в клетку эпидермиса (рис 5). Прорастание зооспоры сопровождается формированием многоядерной плазменной массы (плазмодий), которая покрывается оболочкой и превращается в зооспорангий с длинным горлышком, через которые зооспоры выходят наружу и поражают новые растения. При неблагоприятных условиях зооспоры попарно копулируют, образую двужгутиковую зиготу, которая проникает в клетку хозяина, увеличивается, покрывается плотной бородавчатой оболочкой и превращается в цисту, которая является покоящейся стадией и зимует.
Весной циста прорастает зооспорангием с зооспорами (рис 5).
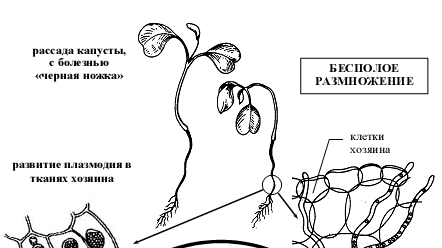
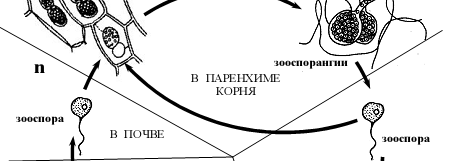
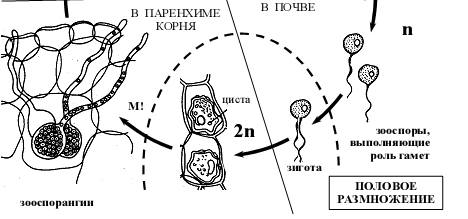
Рисунок 5 - Схема жизненного цикла ольпидиума капустного (Olpidium brassicae), возбудителя «черной ножки» рассады капусты.
- Жизненный цикл Мукора (Mucor mucedo ) .
Для мукора характерны только многоспоровые спорангии. Когда на верхушке спорангиеносца появляется вздутие –будущий спорангий, оно отделяется от спорангиеносца куполообразной перегородкой, вследствие чего спорангиеносец вдается в полость спорангия. Та часть спорангиеносца, которая находится внутри спорангия, называется колонкой. Существование колонки – отличительная особенность бесполого спороношення не только мукора, но и представителей всего его семейства. Зрелые спорангии не отделяются от спорангиеносца. Генетически однородные спорангиоспоры освобождаются в результате расплывания или разрушения оболочки спорангия. Споры дают начало новым мицелиям (рис 6).
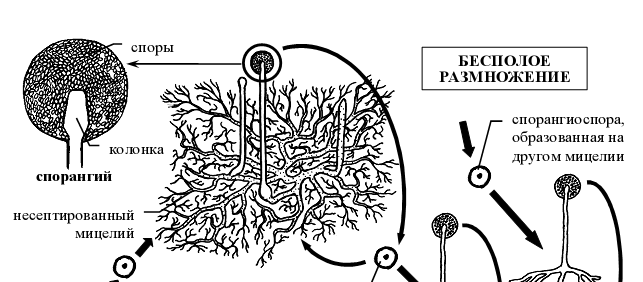
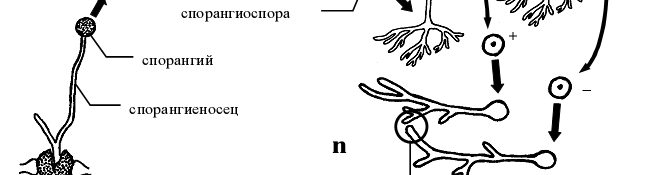
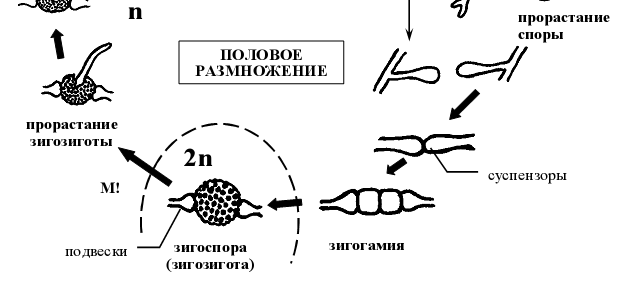
Рисунок 6 - Схема жизненного цикла представителей рода мукор (Mucor mucedo )
Половой процесс представляет собой типичную зигогамию (га- метангиогамия, при которой происходит слияние неспециализированных одно- или многоядерных участков несептированного мицелия). Началом полового процесса служит столкновение кончиков обычно коротких гиф мицелия, несколько вздувающихся на концах и отделяющихся перегородкой от несущей гифы. Перегородка между образовавшимися конечными клетками исчезает, и они сливаются. Заканчивается половой процесс попарным слиянием ядер (так называемая множественная кариогамия). Зигота после состояния покоя прорастает, образуя короткую гифу со спорангием на конце (так называемый зачаточный спорангий). Ее прорастанию обычно предшествует редукционное деление диплоидных ядер, что обуславливает генетическую разнородность спор, образованных в зачаточном спорангии. Необходимо отметить, что двуядерное состояние зиготы сохраняется весь период покоя, а кариогамии происходят непосредственно перед прорастанием [1].
- Жизненный цикл схизосахаромицетов и сахаромицетов см. вопрос 3.
- Жизненный цикл спорыньи ( Claviceps purpurea) .
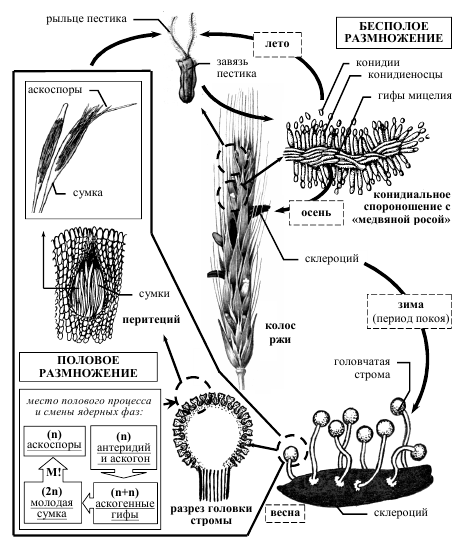
Рисунок 7 – Схема жизненного цикла спорыньи пурпурной Claviceps purpurea.
Склероции зимуют в почве, куда они попадают при уборке урожая с культурных злаков или дикорастущих злаков, встречающихся по краям полей. Весной склероции прорастают несколькими головчатыми стромами. Их количество и размеры зависят от размеров склероциев. Стромы обычно красноватого цвета. По периферии головок стромы закладываются многочисленные камеры, каждая из которых содержит многоядерный аскогон без трихогины и расположенные на той же гифе многоядерные антеридии (спорынья гомоталлична). После плазмогамии развиваются аскогенные гифы, а на них сумки, содержащие по восемь аскоспор. Одновременно образуется тонкий перидий, отделяющий перитеций от ткани стромы – образуются перитеции. После выбрасывания из перитециев аскоспоры разносятся ветром и попадают на растения. Аскоспоры спорыньи заражают злаки в период цветения. Если аскоспора попадает на цветковые чешуи, ее ростковая трубка не способна через них проникнуть и заражения не происходит. Если же цветковые чешуи открыты, аскоспоры попадают на рыльце пестика или в нектар, прорастают, и мицелий достигает завязи. Необходимо отметить, что до цветения культурных злаков спорынья с успехом поражает дикорастущие и образует на них конидиальное спороношение и даже склероции, дожидаясь массового цветения основного хозяина. Через несколько дней после заражения на растениях развивается конидиальная стадия гриба, имеющая собственное название сфацелия (Sphacelia). В завязи образуется плотная масса мицелия, покрытая слоем конидиеносцев, продуцирующих огромное количество мелких конидий, погруженных в капли «медвяной росы». Медвяная роса –сладковатая жидкость, имеющая неприятный запах и содержащая большое количество сахаров. «Медвяная роса» играет существенную роль в распространении конидий гриба. Привлеченные ею насекомые переносят конидии на здоровые растения. Конидии могут распространяться и каплями дождя, при трении колосьев друг о друга под действием ветра, а после высыхания капель «медвяной росы» они могут переноситься ветром. В пораженных цветках ткани завязи полностью разрушаются и замещаются мицелием гриба [2].
Ко времени окончания цветения злаков и созревания зерна мицелий спорыньи уплотняется и превращается в склероций. Склероции развиваются медленно. Сначала они желтовато-коричневые, затем приобретают серо-фиолетовую или черно-фиолетовую окраску. Полное их созревание приурочено ко времени спелости зерна.
- Жизненный цикл монилинии ( Monilinia fructigena )
В цикле их развития всегда наблюдается конидиальная стадия типа монилия – Monilia – подушечки коротких конидиеносцев, на которых образуются длинные, часто ветвящиеся цепочки конидий (рис 8). М. fructigena вызывает плодовую гниль яблок и груш. Гриб заражает плоды с поврежденной кожицей. Инфекция часто переносится жуком-казаркой.
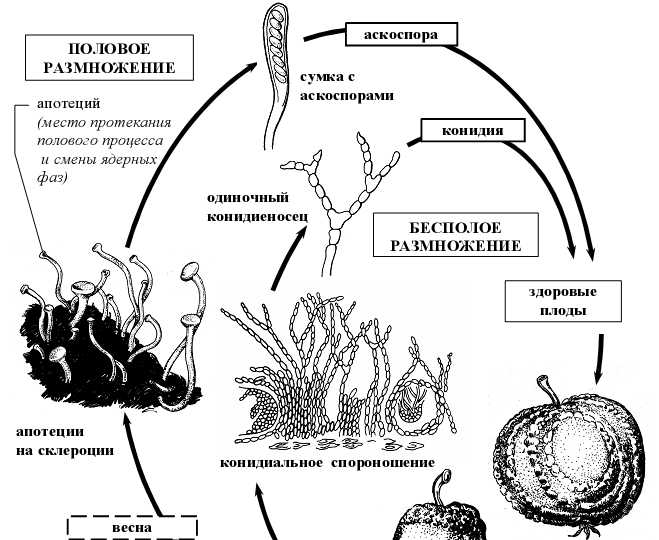
Рисунок 8 – Схема жизненного цикла монилинии (Monilinia fructigena)
Питаясь пораженными плодами, жук при откладывании яиц переносит конидии паразита в здоровый плод на лапках, брюшке и в экскрементах. Конидии гриба прорастают одновременно с развитием личинок жука, которые питаются зараженной грибом мякотью плода. На пораженных плодах образуются пятна отмершей ткани, а на них концентрическими кольцами развиваются желтоватые подушечки конидиального спороношения гриба. Зараженные плоды опадают с дерева и служат источником инфекции. Плоды, оставшиеся на дереве, превращаются в склероции. В пораженных плодах формируются полые шаровидные склероции, снаружи и внутри покрытые черной корой. Они располагаются под эпидермисом пораженных яблок, приобретающих в связи с этим черную окраску. Склероции зимуют, а весной на них снова развивается конидиальное спороношение гриба. Апотеции у этого вида образуются редко [1].
- Жизненный цикл базидиомикотовых на примере агариковых.
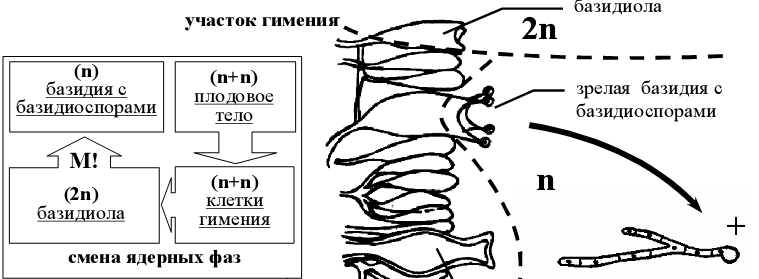
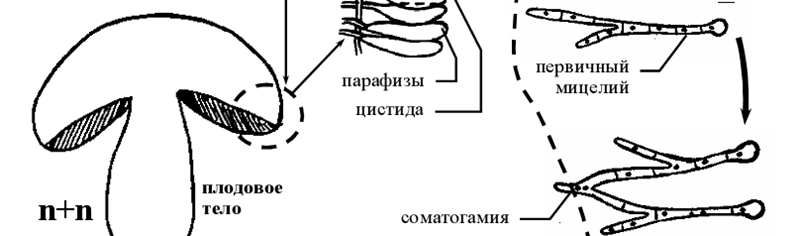
Рисунок 9 – схема жизненного цикла базидиальных грибов
В цикле развития базидиальных грибов наблюдается чередование гаплоидной, дикарионтичной и диплоидной ядерных фаз. В отличие от сумчатых грибов, преобладает дикарионтичная фаза: вторичный мицелий, плодовые тела (рис 9); гаплоидная фаза короткая: базидиоспоры и первичный мицелий, выросший из них; диплоидная фаза представлена молодой базидией (базидиолой).
Созревающие базидии, базидиоспоры и вырастающий из них первичный мицелий – гаплоидные [1].
Первичный мицелий существует непродолжительный период времени, затем происходит половой процесс в виде слияния двух гаплоидных вегетативных клеток мицелия (соматогамия). Среди агариковых грибов преобладают гетероталличные (раздельнополые) виды, у которых для слияния двух клеток при образовании дикариотичного мицелия должны встретиться гифы, выросшие из базидиоспор разных знаков.
При соматогамии происходит слияние цитоплазмы, а ядра объединяются в пары – дикарионы, которые затем синхронно делятся.
Вторичный дикарионтичный мицелий может существовать длительное время, интенсивно разрастается в субстрате и образует плодовые тела (рис 9).
Плодовые тела агариковых грибов, так же как грибница, пронизывающая субстрат, состоят из дикарионтичных гиф. На концах дикарионтичных гиф гимениального слоя происходит формирование базидий. В развивающейся базидии завершается половой процесс: сливаются ядра дикариона (молодая базидия – базидиола – диплоидна), а затем происходит редукционное деление, в результате которого образуется четыре гаплоидных ядра. Каждое ядро по специальному каналу (стеригме) переходит в вырост базидии, где после закрытия канала формируются обособленные базидиоспоры [2].
- Жизненный цикл пыльной головни (Ustilago tritici)
Возбудитель пыльной головни пшеницы ( U. tritici ) разрушает зерна и колоски злака (в колосе, вышедшем из листового влагалища, сохраняются лишь стержень и разрушенные боковые колоски), превращая остальные части колоса в темную пылящую массу телиоспор.
У данного представителя телиоспоры не являются зимующей (покоящейся) стадией, а служат для расселения гриба и прорастают без периода покоя. Они легко переносятся ветром на цветущие в это время колосья, заражая завязи, и, попав на рыльце пестика питающего растения, прорастают четырех-клеточной базидией. При прорастании двухъядерной телиоспоры происходит кариогамия, а затем – редукционное деление с формированием гаплоидных ядер. Образования базидиоспор у возбудителя пыльной головни пшеницы не происходит, а дикарион возникает путем перехода ядер из одной клетки базидии в другую либо путем копуляции соседних клеток базидии или клеток соседних базидий. Из клетки базидии, ставшей двухъядерной, вырастает дикариотический мицелий, который по пыльцевой трубке проникает в завязь к зародышу семени. В виде мицелия паразит и зимует.
Однако по мере дальнейшего развития растения одновременно трогается в рост и мицелий гриба, проникающий в конус нарастания и растущий вместе со стеблем. Заканчивается вегетация гриба образованием телиоспор, после чего вместо нормального колоса остается темная обугленная ось соцветия [1].
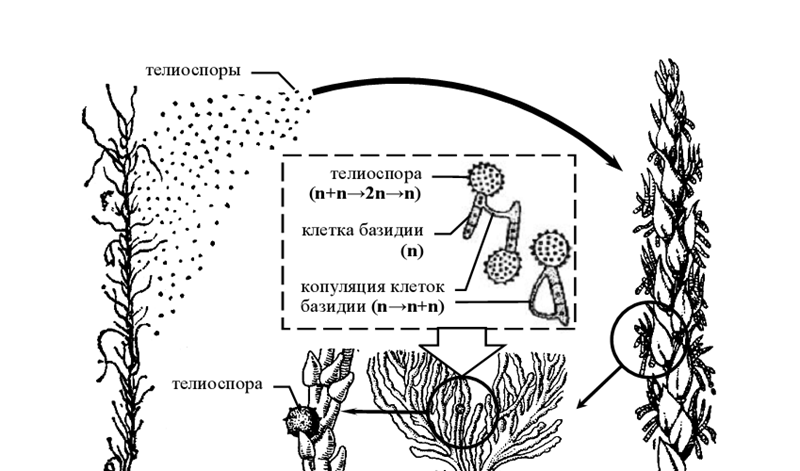
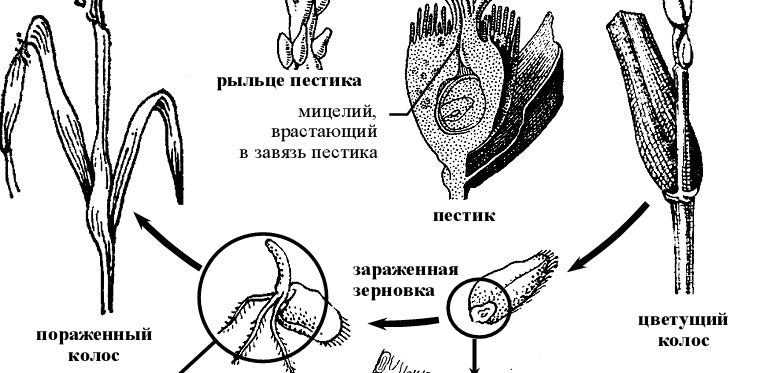
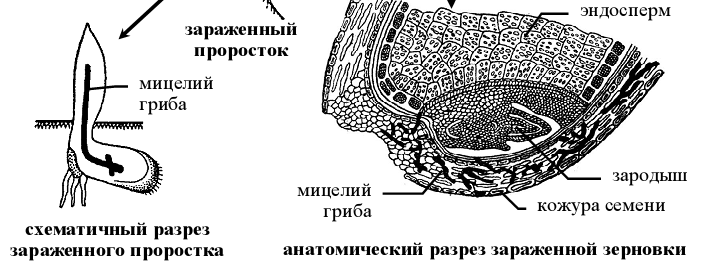
Рисунок 10 – Схема жизненного цикла пыльной головни пшеницы ( Ustilago tritici).
- Жизненный цикл твердой головни ( Тilletia caries)
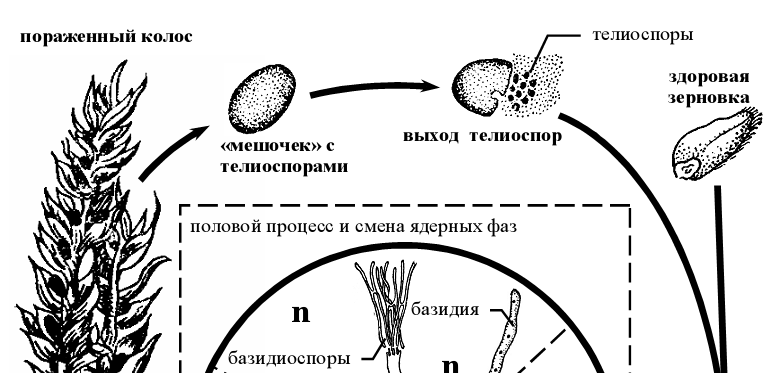
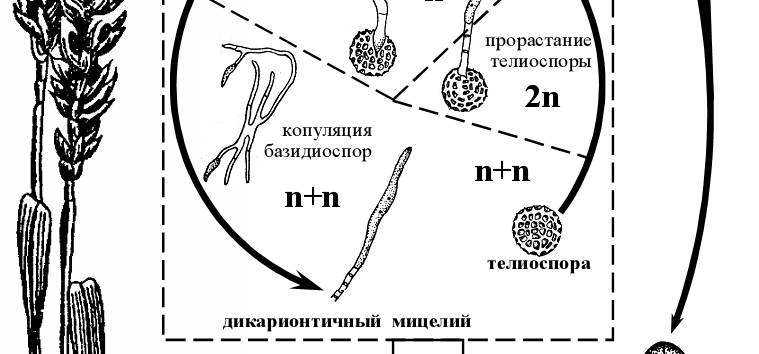
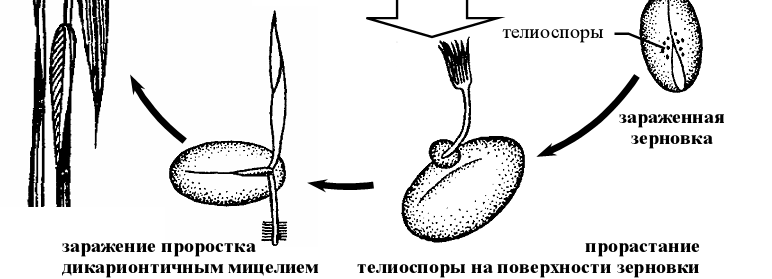
Рисунок 11 - Схема жизненного цикла возбудителя твердой головни пшеницы (Тilletia caries ).
Покоящейся стадией данного гриба-паразита являются телиоспоры, которые прикрепляются к зерновкам в конце периода вегетации растения либо при обмолоте зерновых. Вместе с зерновкой двухъядерные телиоспоры попадают в почву, где весной прорастают в базидию с длинными нитевидными базидиоспорами на вершине. Перед формированием базидий в телиоспорах происходит слияние ядер и редукционное деление. Палочковидные гаплоидные базидиоспоры попарно копулируют между собой через копуляционные мостики. Из образующихся двухъядерных клеток развивается дикарионтичный мицелий, проникающий в конус нарастания проростка, затем по мере развития растения – в завязь пестиков, где поглощает питательные вещества завязи. Мицелий разрастается и вскоре распадается на телиоспоры. Вместо зерновки образуется головневый мешочек, заполненный телиоспорами. При раздавливании околоплодника зерновки становится видна мажущаяся, жирная на ощупь, темно-коричневая масса спор с селедочным запахом, за что болезнь еще называют мокрой или вонючей головней [2].
- Жизненный цикл Пункции злаковой ( Puccinia graminis)
Развитие гриба начинается с прорастания зимующих на соломе телейтоспор и образования базидии с базидиоспорами. Этому предшествует кариогамия (образование диплоидного ядра), а затем редукционное деление. Гаплоидные ядра по одному попадают в каждую клетку базидии, и каждая из них формирует одну базидиоспору, куда и переходят гаплоидные ядра [1].
Базидиоспоры – маленькие тонкостенные клетки, сидящие на тонких стеригмах. При созревании они отбрасываются на некоторое расстояние и затем распространяются воздушными течениями. Для дальнейшего развития базидиоспоры должны попасть на листья барбариса, где протекает весенняя фаза развития возбудителя стеблевой ржавчины. После проникновения ростка базидиоспоры в ткань листа сначала появляются оранжевые пятна, образованные гипертрофированной тканью листа, пронизанной по межклетникам мицелием гриба с гаусториями, внедрившимися в клетки (рис 12).
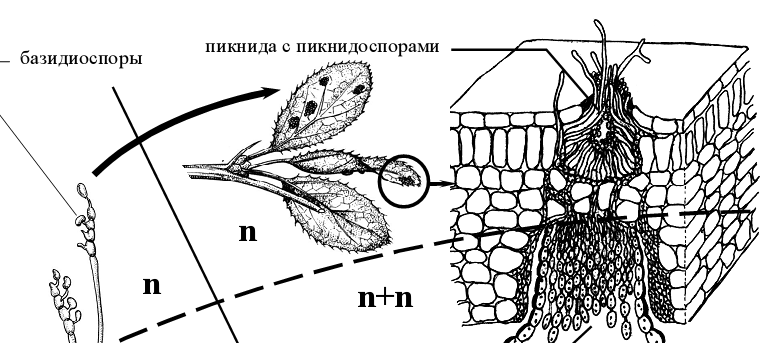
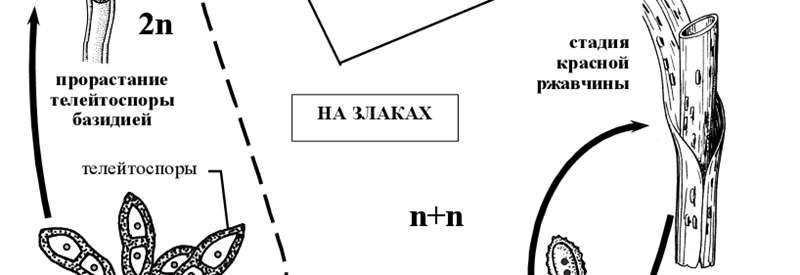
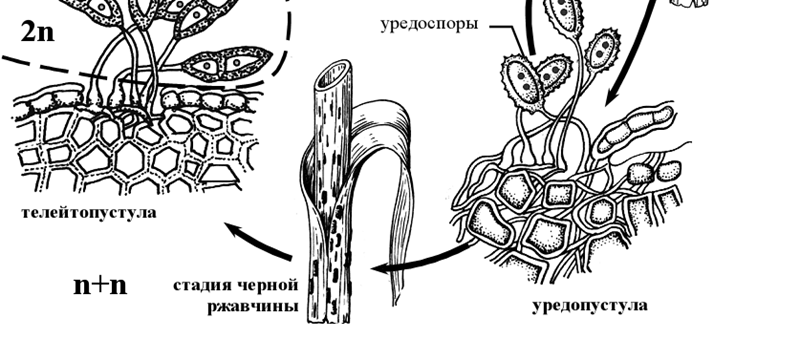
Рисунок 12 – Схема жизненного цикла стеблевой ржавчины злаков (Puccinia graminis)
Гифы состоят из одноядерных клеток с оранжевыми каплями масла и принадлежат к гаплоидной стадии развития гриба. На таком мицелии закладываются кувшиновидные спороношения – спермогонии (пикниды, пикнии), в которых развиваются спермации (пикнидоспоры, пикниоспоры). Спермации вместе с выделяющейся сладковатой жидкостью выносятся на
поверхность листа и переносятся насекомыми на другие спермогонии (сперматизация). Там они прорастают в гаплоидный мицелий, и его гифы, срастаясь с гифами другого полового знака, образуют дикариотический мицелий. Такой мицелий прорастает через толщу листа на его нижнюю сторону и образует зачатки эцидиального дикарионтического спороношения. Эцидии (эции) первоначально дифференцируются на две части: стерильную, сложенную из крупных, бедных содержимым пузыревидных клеток, и плодущую, сложенную из гиф, богатых цитоплазмой. На гифах дифференцируются базальные клетки, которые начинают отшнуровывать на своем конце цепочку материнских клеток эцидиоспор. Материнская клетка эцидиоспоры делится (каждая) на две двухъядерные неравные по объему клетки. У концевой, более крупной клетки, вырабатывается толстая оболочка, и она превращается в эцидиоспору (эциоспоры), промежуточная клетка остается тонкостенной и позднее разрушается [2].
Округлые двухъядерные эцидиоспоры по мере созревания выпадают из урновидного эцидия, и распространяются по воздуху. Заражать барбарис они не могут и для дальнейшего развития должны попасть на листья или стебли злаков. На злаках эцидиоспоры прорастают мицелием, который проникает через устьица в ткани, где формируется межклеточный дикарионтичный мицелий с гаусториями. На таком двухъядерном мицелии развивается летнее спороношение гриба – уредопустулы с уредоспорами. Каждая уредоспора сидит на клетке-ножке. Под давлением уредоспороношений эпидермис листа или стебля разрывается, обнажая желтые, бурые, кирпично-красные или оранжевые подушечки уредоспор – то, что называется красной ржавчиной.
Уредоспоры рассеиваются в воздухе и могут снова заражать злаки. Прорастая, они развивают дикарионтический мицелий, проникающий через устьица в ткани растений. Через несколько дней снова образуются уредоспоры, вызывающие новые заражения растений. Таких новых поколений уредоспор за лето может быть до десяти. Чем быстрее проходит период от заражения до возникновения нового поколения уредоспор, тем сильнее развивается на злаках ржавчина.
По мере истощения питательного субстрата (растения) на том же дикариотическом мицелии возникают новые спороношения –телейтопустулы (телии) с телейтоспорами, или зимними спорами. У возбудителя стеблевой ржавчины они двухклеточные, сидят на ножке, каждая из клеток первоначально имеет по два ядра, позднее сливающихся в одно диплоидное ядро. Оболочки телейтоспор толстые, темно-бурые, с одной ростковой порой в каждой клетке. При поражении злаков стеблевой ржавчиной к концу вегетации подушечки телейтоспор покрывают стебли и листовые влагалища черными продольными полосами: наступает стадия черной ржавчины. Телейтоспоры возбудителя стеблевой ржавчины служат для перезимовки гриба, прорастая лишь после периода покоя. Зимуют они на стерне или соломе (в скирдах), а на диких злаках также на стоящих зимующих стеблях. Весной каждая клетка телейтоспоры прорастает четырехклеточной базидией [1].
- Жизненный цикл Пиннулярии ( Pinnularia viridis)
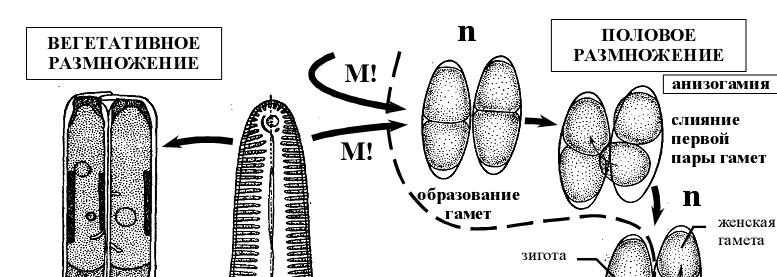
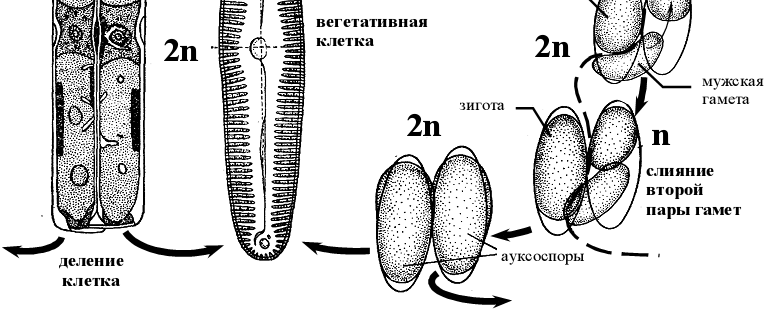
Рисунок 13 – Схемы жизненного цикла пиннулярии Pinnularia viridis.
В ходе последовательных мейотического и митотического делений в каждой материнской клетке образуется по одной подвижной (мужской) и одной неподвижной (женской) гамете. Подвижные гаметы передвигаются к неподвижным и сливаются с ними. Во втором случае в одной клетке обе гаметы неподвижные, в другой – обе подвижные, переходящие в клетку с неподвижными гаметами [6].
2. Жизненный цикл хлорококка (Chlorococcum)
При бесполом размножении в материнской клетке образуется от 8 до 32 двужгутиковых зооспор, которые освобождаются через разрыв оболочки. Через некоторое время они теряют жгутики, одеваются плотной оболочкой и превращаются в неподвижные вегетирующие клетки хлорококкума, постепенно дорастающие до размеров материнской. Половой процесс происходит путем слияния двужгутиковых изогамет, похожих на зооспоры. Зигота прорастает после периода покоя. Редукция зиготическая [6].
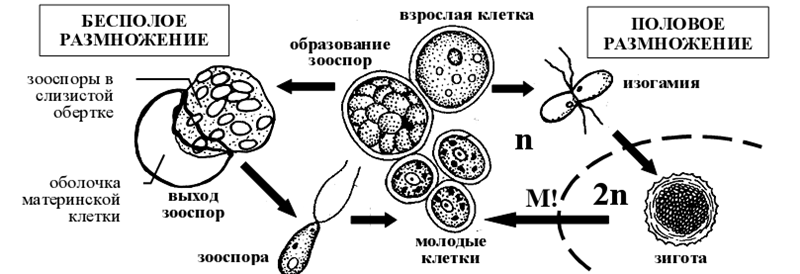
Рисунок 14 – Схема жизненного цикла представителей рода хлорококкум ( Chlorococcum).
- Жизненный цикл фитофторы инфекционной смотреть выше.
Порядок заключения коллективного договора коллективного договора Условия коллективного договора носят обязательный характер. Коллективный договор как институт трудового права это совокупность правовых норм которые регулируют трудовые и социально-экономические отношения между наемными работниками и работодателями определяют порядок разработки заключения и исполнения трудового договора комплексно регулируют различные вопросы которые касаются разных аспектов трудовых правоотношений.
Рекомендуем ознакомится: http://5fan.ru