Строение акул
Большинство современных акул имеют удлиненную форму тела. Исключением являются акулы отрядов морских ангелов (Squatiniformes) и пилоносов (Pristiophoriformes), имеющие сжатую в дорсовентральном направлении скатообразную форму тела. В отличие от скатов у акул жаберные щели открываются по бокам головы (у скатов — на ее вентральной стороне). Жаберных щелей у акул обычно 5, но у представителей древнего отряда Hexanchiformes и пилоносов (Pliotrema) — 6 или 7. Жаберная крышка отсутствует. Передняя жаберная щель редуцирована и у большинства акул представлена небольшим отверстием — брызгальцем (рис. 1).
Тело акул заканчивается мощным хвостовым стеблем, несущим хвостовой плавник, чаще всего — типично гетероцеркальным (верхняя лопасть заметно больше нижней). У большинства акул семейства Lamnidae хвостовой плавник почти гомоцеркальный (лопасти почти равны). На спинной стороне тела расположены один или два спинных плавника, иногда перед ними есть колючка, или шип (Heterodontiformes, Squalidae). У всех акул, кроме колючих (Squaliformes), пилоносов и скватинид, имеется непарный анальный плавник. Все они имеют две пары парных плавников — грудные и брюшные. У ряда видов на хвостовом стебле есть кили (рис. 1, а).
Ротовое отверстие у акул большое, расположено на нижней стороне головы (кроме плащеносной и мегапастной акул). Челюсти вооружены крупными зубами. На голове расположены глаза и ноздри. Последние чаще всего занимают также вентральное положение. Голова акулы-молота имеет своеобразную форму: боковые выросты придают ей форму молота или лопаты, на растральных краях которых расположены ноздри. Глаза у акул этого семейства лежат на боковых плоскостях «молотов».
У всех акул имеется клоака. Самцы всех видов отличаются от самок наличием птеригоподиев, или наружных половых органов, хорошо различимых невооруженным глазом уже у эмбрионов длиной 55—60 мм.
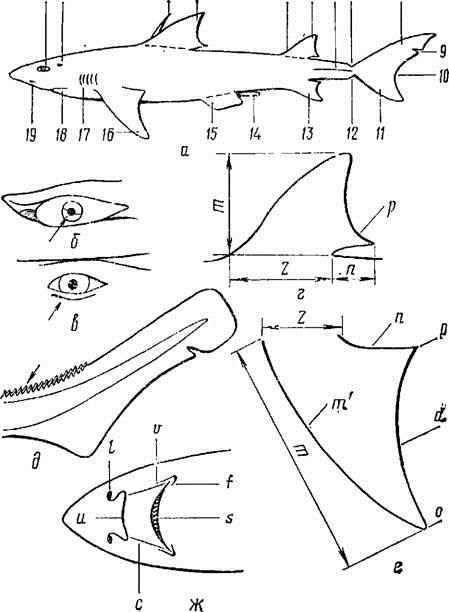
Рис. 1. Внешнее строение акулы:
а — внешний вид: 1 — глаз; 2 — брызгальце: 3— шип, или колючка, спинных плавников; 4 — первый спинной плавник; 5 — второй спинной плавник; 6 — киль хвостового стебля; 7 — верхняя предхвостовая ямка; 8 — верхняя лопасть хвостового плавника; 9 — выемка верхней лопасти хвостового плавника; 10 — выемка хвостового плавника; И — нижняя лопасть хвостового плавника; 12 — нижняя предхвостовая ямка; 13 — анальный плавник; 14 — птеригоподии самца; 15 — брюшные плавники; 16 — грудные плавники; 17 — жаберные щели; 18 — рот; 19 — ноздря; б — мигательная перепонка глаза акулы; в — складка нижнего века акулы; г — непарный плавник: z — длина основания; т — высота; и — длина свободного конца плавника; р— свободный конец плавника, д — гребень из плакоидных чешуй на хвостовом стебле акулы; е — парный плавник: z — длина основания; т — высота плавника, т' — внешний край плавника; п — передний внутренний край; р — угол внутренний; d — задний внутренний край; о — вершина плавника; ж — вид головы акулы снизу: I — ноздри; v — верхняя околоротовая бороздка; / — нижняя околоротовая бороздка; s — рот; с — ротоносовая бороздка; и — носовой клапан.
Внутреннее строение акулы
Кожа. Кожа акул состоит из многослойного эпидермиса и подстилающего его соединительнотканного слоя — кориума. В эпидермисе расположены многочисленные одноклеточные железы, слизистый секрет которых равномерно покрывает все тело акулы. Пигментные клетки расположены как в нижних слоях эпидермиса, так и в кориуме.
В коже акул расположены (и постоянно образуются) плакоидные чешуи, представляющие собой шипики, оболочка которых состоит из дентина. Вершина чешуйки покрыта плотным слоем эмали. Внутри чешуйки имеется полость. заполненная рыхлой тканью, насыщенной кровеносными сосудами. Чешуи, как и зубы акул, имеют самую разнообразную форму.
Плакоидные чешуи больших размеров, расположенные на челюстях, видоизменяются в зубы акул. Они отличаются от собственно чешуй кроме размеров и формы значительно увеличенным слоем эмали и дентина и уменьшением полости с рыхлой тканью.
Скелет. У всех современных акул в скелете отсутствуют кости; внутренний скелет полностью хрящевой. У крупных особей происходит значительное обызвествление хрящей, особенно позвоночника.
Осевой скелет акул представлен позвоночным столбом, или позвоночником, разделяющимся на туловищный (прекаудальный) и хвостовой отделы по форме тел позвонков. Позвонки акул амфицельные или двояковогнутые. В центре позвонка находится канал, по которому проходит хорда. Хорда имеет вид бус и проходит по всему позвоночнику, расширяясь в полостях между позвонками и сужаясь внутри них. Хорда у акул не несет опорной функции.
На дорсальных (спинных) позвонках расположены верхние дуги, в месте слияния которых образуются короткие остистые отростки. Между верхними дугами расположены вставочные пластинки, образующие вместе с первыми стенки спинномозгового канала, в котором лежит спинной мозг.
На прекаудальных (туловищных) позвонках (рис. 2, б) имеются короткие поперечные отростки, образованные нижними дугами, к которым причленяются очень короткие ребра. Нижние дуги хвостовых (рис. 2, а) позвонков смыкаются друг с другом, образуя гемальный канал, по которому проходят хвостовая артерия и вена.
Череп акул, как и других позвоночных, разделяется на два отдела — мозговой, или нейрокраниум, и висцеральный.
Мозговой череп акул представляет собой сплошную хрящевую коробку, со всех сторон окружающую головной мозг. В верхней части черепа есть небольшое отверстие, затянутое соединительной тканью. Передний конец мозгового черепа вытянут вперед в рост- рум (кроме плащеносной акулы). У основания последнего с черепом срастаются парные обонятельные капсулы, а в заднюю часть боковых стенок врастают слуховые капсулы. Мозговой череп акул платибазального типа. Основание его широкое. Стенки глазниц разобщены, между ними лежит головной мозг.
Висцеральный череп (рис. 2, в) состоит из преобразованных жаберных дуг. Челюстные дуги образованы парными хрящами; функцию верхней челюсти несет небно-квадратный хрящ, образующий челюстной сустав с меккелевым хрящом (нижней челюстью).
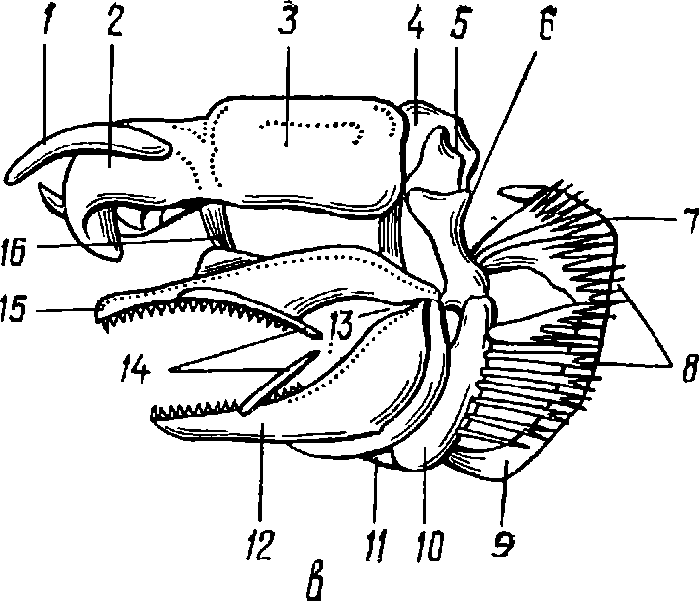
Рис. 2. Элементы скелета акулы
а — хвостовой позвонок; б — туловищный позвонок; в — череп: 1 — рострум; 2— обонятельная капсула; 3— глазница; 4 — слуховой отдел; 5 — затылочный отдел (1—5 — мозговой череп, или нейрокраниум); 6 — гиомандибуляр; 7 и 9— жаберные дуги; 8 — жаберные лучи; 10 — гиоид; 11 —копула подъязычной дуги; 12 — меккелев хрящ; 13 — челюстной сустав; 14 — губные хрящи; 15 — небно-квадратный хрящ; 16 — связки
На обеих челюстях есть зубы. У многих акул есть 1—2 пары мелких губных хрящей, расположенных спереди челюстной дуги.
Подъязычная, или гиоидная, дуга висцерального черепа лежит сразу за челюстной. Крупный парный верхний хрящ — гиомандибуляр — подвижно прикреплен к слуховому отделу мозгового черепа, а его нижний конец подвижно сочленен с челюстной дугой и гиоидом. Правый и левый гиоиды соединяются друг с другом через мелкий хрящ — копулу. Такое соединение челюстей с мозговым черепом называется гиостилией и характерно для сравнительно новых видов акул. Для древних видов акул (Hexanchiformes) характерна амфистилия: челюстная дуга прикрепляется к мозговому черепу через подвесок, и, кроме того, отросток небно-квадратного хряща сочленяется с основанием мозгового черепа.
За подъязычной дугой расположены 5—7 пар жаберных дуг, разделенных жаберными щелями. Каждая дуга состоит из подвижно сочленяющихся друг с другом четырех парных элементов и замыкающей дугу снизу непарной копулы. Верхние элементы дуги соединяются с позвоночником.
Скелет непарных плавников акул образован палочковидными хрящами — радиалиями, погруженными в мускулатуру и достигающими основания плавника. Собственно лопасть плавника поддерживается эластотрихиями — тонкими лучами из рогоподобного вещества.
Пояс передних конечностей акул имеет вид хрящевой дуги, лежащей в толще мускулатуры за жабрами, а иногда частично под ними. Он не связан со скелетом позвоночника и черепа. Часть пояса, лежащая выше сочлененного выроста, к которому прикрепляется основание грудных плавников, называется лопаточным отделом, а лежащая ниже — каракоидным.
Тазовый пояс, или пояс брюшных плавников, акул образован хрящевой пластиной, лежащей в мускулатуре брюшной стенки непосредственно перед клоакой. К ее боковым поверхностям причленяется скелет плавников, состоящий из одного удлиненного базального элемента, к наружной поверхности которого прикрепляются радиальные хрящи брюшного плавника. У самцов акул базальный элемент удлинен и служит скелетной основой птеригоподия.
Мышечная система. Соматическая мускулатура акул отчетливо сегментарна и состоит из мышечных сегментов — миомеров. Последние разделены соединительнотканными перегородками — миосептами. На отдельных участках тела метамерия нарушается: перестройка миомеров или их участков приводит к дифференцировке отдельных мышц. Так обособляются глазные, наджаберные и поджаберные мышцы и мышцы парных плавников.
Висцеральная мускулатура представляет собой слои гладких мышц, окружающих пищевод. В области челюстей и жабр у акул вместо гладких мышечных волокон висцеральной мускулатуры находятся поперечнополосатые мышцы, которые формируют сложные дифференцированные группы мышц, управляющие движениями челюстей и жаберных дуг.
Характерной чертой мускулатуры акул является ее автономность. Мускулы акул сохраняют способность сокращаться и при нарушении связи с центральной нервной системой. Этим объясняется хорошо известная живучесть акул, плавающих некоторое время без внутренностей. Особенно долго сохраняют способность к сокращению мышцы челюстей, что необходимо помнить при промысле и изучении акул, так как часто -уже мертвые акулы наносят тяжелые травмы своими зубами.
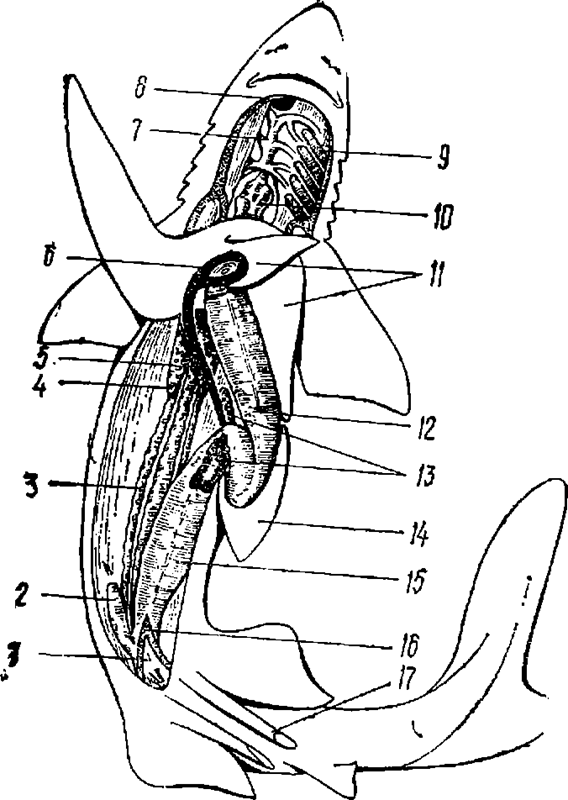
Рис. 3. Внутреннее строение половозрелого самца акулы.
1 — клоака; 2 — ректальная железа; 3 — семяпровод; 4 — семенник; 5 — желчный проток; 6 — желчный пузырь; 7 —жаберные артерии; 8 — щитовидная железа; 9 — жабры; 10— сердце; 11— лопасти печени; 12 — желудок ; 13 — поджелудочная железа; 14 — селезенка; 15 — толстая кишка и спиральный клапан; 16 — прямая кишка; 17 — птеригоподии (совокупительные органы самца)
Пищеварительная система. Захват и частичная механическая переработка пищи производятся челюстями акул, снабженными сложной системой мускулатуры и мощными совершенными зубами. Это особенно характерно для акул-бентофагов (Heteroclontiformes, Triakidae), зубы которых образуют своего рода «терку», хорошо перетирающую раковины моллюсков и панцири ракообразных.
На дне ротовой полости акул находится небольшая слизистая складка — язык, движения которого обусловливаются движениями подъязычных дуг. Ротовая полость переходит в глотку, стенки которой пронизаны жаберными отверстиями. Прохождению пищи через них препятствуют хрящевые жаберные тычинки. Короткий пищевод без заметных границ переходит в объемистый желудок V-об- разной формы.
В передней (кардинальной) части желудка (рис. 3) происходит длительное (до 5 сут и более) переваривание пищи. Затем она поступает в более узкую (пилорическую) часть желудка. За желудком начинается кишечник, отчетливо разделяющийся на три отдела. Очень короткая тонкая кишка отделена от желудка кольцевым утолщением мускулатуры (сфипктром). В тонкую кишку открываются протоки поджелудочной железы и желчный проток. Внутри широкой и длинной толстой кишки расположен вырост ее стенки — спиральный клапан, образующий 10—15 витков, что значительно увеличивает внутреннюю поверхность кишечника. Спиральный клапан плавно переходит в короткую прямую кишку, открывающуюся в клоаку.
Детали строения и соотношение отделов кишечника у различных видов акул несколько варьируют, что обусловлено особенностями их питания.
Трехлопастная печень акул имеет большие размеры, ее масса составляет около 10%, а у отдельных видов акул — около 20% массы тела. Печень акул накапливает огромные количества жира (до 70 % массы), вследствие чего она выполняет роль не только депо энергетических ресурсов, но и гидростатического органа, повышающего плавучесть тела акулы, лишенного плавательного пузыря.
Жир печени акул содержит большое количество сквалена (более 50 % у отдельных кошачьих акул) и витаминов. Так, в печени черноморского катрана содержание жира достигает 80 % массы печени, а витамина А в 1 г этого жира — 830 ИЕ.
Органы дыхания. Стенки глотки акул пронизаны открывающимися наружу жаберными щелями. К подъязычной дуге и жаберным дугам прикрепляются межжаберные перегородки, края которых образуют кожистую складку над следующей жаберной щелью. По бокам этих перегородок расположены жаберные лепестки эктодермального происхождения. Совокупность жаберных лепестков на обеих сторонах перегородки образует жабры.
У большинства акул между челюстной и подъязычной дугами сохраняется остаток жаберной щели в виде отверстия, лежащего за глазом, — брызгальце, которое ведет в начальную часть глотки. Через него и ротовую полость в глотку акул всасывается вода, которая затем выбрасывается через жаберные щели, омывая жаберные лепестки и насыщая кровь акул кислородом. В отличие от скатов у акул функциональное значение брызгальца очень мало (за исключением, вероятно, очень малого числа видов).
Кровеносная система. У акул и других хрящевых рыб имеется только один круг кровообращения, и в сердце поступает только венозная кровь.
Сердце у акул двухкамерное, состоит из предсердия и желудочка. Кроме этих основных отделов имеются тонкостенный венозный синус, или венозная пазуха, и артериальный конус. Последний представляет собой расширенное начало брюшной аорты, в котором, как и в остальных отделах сердца, стенки образованы поперечнополосатой мускулатурой, а не гладкими мышечными волокнами, как стенки кровеносных сосудов. Внутри артериального конуса находятся клапаны, препятствующие обратному току крови.
От артериального конуса отходит брюшная аорта, распадающаяся на парные приносящие жаберные артерии. От первой из них ответвляется общая сонная артерия, снабжающая кровью голову. Выносящие жаберные артерии обеих сторон сливаются, образуя лежащую под позвоночником спинную аорту. От последней отходят парные подключичные артерии, идущие к грудным плавникам и жаберному аппарату. В туловищной области от спинной аорты отходит ряд парных и непарных артерий, идущих к плавникам и внутренним органам. Далее хвостовая аорта переходит в хвостовую» артерию.
Венозная кровь возвращается в сердце по венам, более широким и тонкостенным, чем артерии. Хвостовая вена входит в полость тела и разделяется на правую и левую воротные вены почек. В почках вены распадаются на капилляры, образуя воротную систему почек. Почечные капилляры вновь сливаются в вены, образуя парные задние кардинальные вены. По бокам тела акул идут парные боковые вены, которые в области сердца сливаются с соответствующими подключичными венами. Кровь из головы акул собирается в парные передние кардинальные и парные нижние яремные вены. Последние и задние кардинальные вены сливаются в парные кювьеровы протоки, впадающие в венозный синус сердца. В печени акул также образуется воротная система (система печени), после которой через печеночную вену кровь попадает в венозный синус.
У акул и хрящевых рыб имеется хорошо оформленная селезенка — крупный компактный орган, лежащий около желудка (см. рис. 3), которая служит депо крови и является кроветворным органом.
Органы выделения. Основными органами выделения у акул являются туловищные, или мезонефрические, почки, лежащие в виде удлиненных тел вдоль позвоночника (см. рис. 3, 4). От них отходят почечные канальца, впадающие у самок -в вольфов канал, выполняющий роль мочеточника, у самцов — в мочеточник, обособленный от нижней части вольфова канала и впадающий в клоаку, как и вольфов канал самок.
Половая система. Парные семенники у самцов акул расположены над печенью вдоль пищевода (рис. 4, а). От них отходят протоки семенных каналов — тонкие семявыносящие каналы впадающие в почечные канальцы передней части почки. Эта часть почки играет роль придатка семенника; ее канальца открываются в вольфов канал, который функционирует у самцов как семяпровод. В самой задней части последнего образуется семенной пузырек (хорошо развитый у половозрелых самцов) — расширение семяпровода. Левый и правый семяпроводы через семенной пузырек выходят в полость мочеполового сосочка. Сюда же выходят полые слепые выросты — семенные мешки (редуцированные мюллеровы каналы).
Самки акул имеют парные яичники (рис. 4,6), расположенные там же, где и семенники у самцов. Вольфов канал у самок выполняет только функцию мочеточника. В отличие от самцов у самок хорошо развиты мюллеровы каналы, каждый из которых лежит вдоль брюшной стороны почки. Передние концы мюллеровых каналов, выполняющие роль яйцеводов, огибают передний конец печени и сливаются, образуя воронку яйцевода.
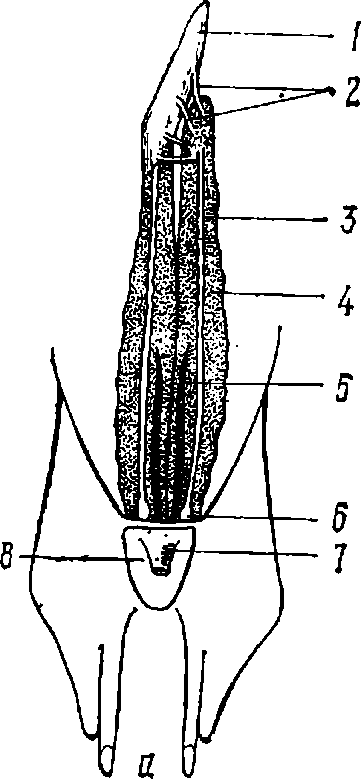
Рис. 4. Половая система самца (а) и самки (б):
1 — семенник; 2 — семявыно- сящие каналы; 3 — семяпровод; 4 — почка; 5 — мочеточник; 6 — семенной мешочек; 7—мочеполовой сосочек; 8 — полость клоаки; 9 — яичник; 10 — общая воронка яйцеводов; 11 — скорлуповая железа; 12 — яйцевод; 13—.расширения яйцеводов, утерусы; 14 — отверстия яйцеводов; 15 — мочевой сосочек.
В области передней части почек на каждом яйцеводе образуется расширение — скорлуповая, или нидементальная, железа, которая имеет наибольшие размеры у половозрелых самок (особенно у яйцекладущих видов). Задняя часть яйцевода яйцеживородящих и живородящих акул заметно расширена и называется маткой или чаще утерусом. У ряда видов акул (и скатов) развивается только один (правый) утерус. Причина подобной неравнозначности парного органа неизвестна. Яйцеводы выходят в клоаку самостоятельными отверстиями.
Центральная нервная система (ЦНС). По сравнению с кругло- ротыми, костистыми рыбами и земноводными центральная нервная система акул (и хрящевых рыб вообще) и особенно головной мозг стоят на более высоком уровне развития и организации.
Относительные размеры головного мозга различных акул и надотряда в целом заметно превышают таковые вышеназванных классов позвоночных и приближаются к относительным размерам мозга большинства птиц и низших млекопитающих (рис. 5). Последнее, по мнению ряда авторов, служит определенным показателем общего уровня организации ЦНС того или иного таксона животных, хотя бы в первом приближении.
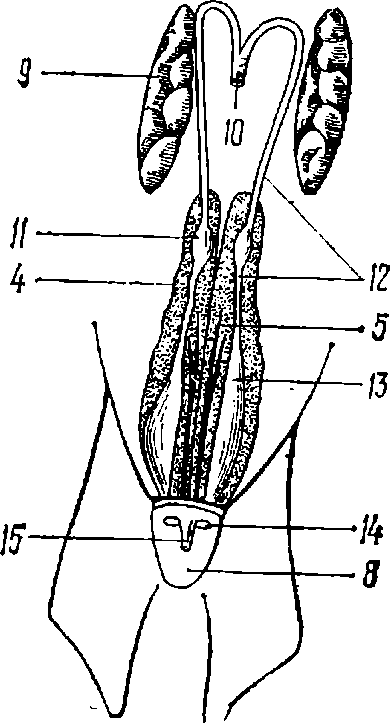
Головной мозг акул представлен всеми основными отделами, характерными для высших позвоночных (рис. 6). Наибольшие размеры имеет передний мозг (как у млекопитающих), масса которого достигает 7г массы всего головного мозга. От переднего мозга к обонятельным мешкам отходят парные обонятельные тракты, соединяющие данный отдел мозга с парными обонятельными луковицами, срастающимися с обонятельным эпителием. Обонятельные луковицы акул выполняют роль первичных обонятельных центров (особенно у акул-бентофагов) и связаны с передним мозгом, где лежат основные обонятельные зоны и ядра; к последнему информация поступает непосредственно из обонятельного эпителия через конечные нервы, которые отсутствуют у костистых рыб. Таким образом, в переднкл мозг акул поступает практически вся информация из ЦНС.
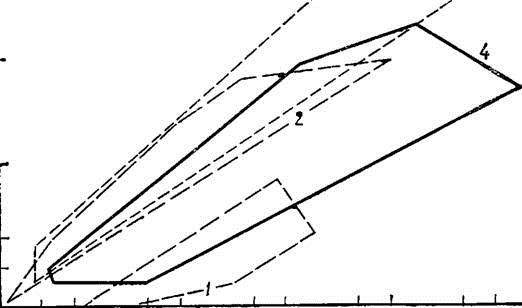
Рис. 5. Полигон энцефализации различных классов позвоночных.
1 — костные рыбы; 2— птицы; 3 — млекопитающие; 4 — акулы (пс данным для 36 видов). По оси абсцисс — масса тела, по ocи ординат — масса головного мозга.
По степени развития обонятельных трактов и луковиц (последних в сопоставлении со зрительными долями) современных акул делят на три группы, характеризующиеся строением мозга и экологией тех или иных видов [65].
За передним мозгом лежит небольшой непарный промежуточный мозг. Средний мозг акул, как и других хрящевых рыб, представлен непарной стволовой частью и парными зрительными долями. Зрительные доли среднего мозга, или зрительный тектум, получают всю первичную информацию от зрения, перерабатывают ее и «передают» далее, в высшие отделы мозга. В стволовой части среднего мозга, а также в продолговатом мозге расположены центры и проводящие пути органов слуха, вкуса и системы органов чувств боковой линии.
Зрительные доли сверху частично прикрыты непарным телом мозжечка. У сравнительно новых видов акул мозжечок покрыт густой сетью продольных и поперечных борозд, что заметно увеличивает площадь его поверхности [67, 190]. Мозжечок является вторым центральным отделом головного мозга акул, также связанный со всеми отделами мозга акул и координирующий их моторные рефлексы, тонус мускулатуры и т, п. Ниже и чуть сзади тела мозжечка расположено более древнее его образование — парные ушки, хорошо выраженные у подавляющего большинства современных акул.
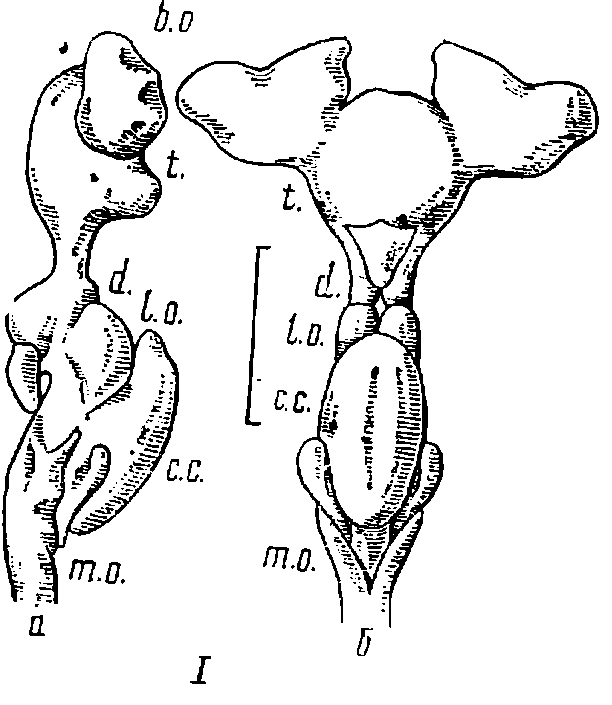
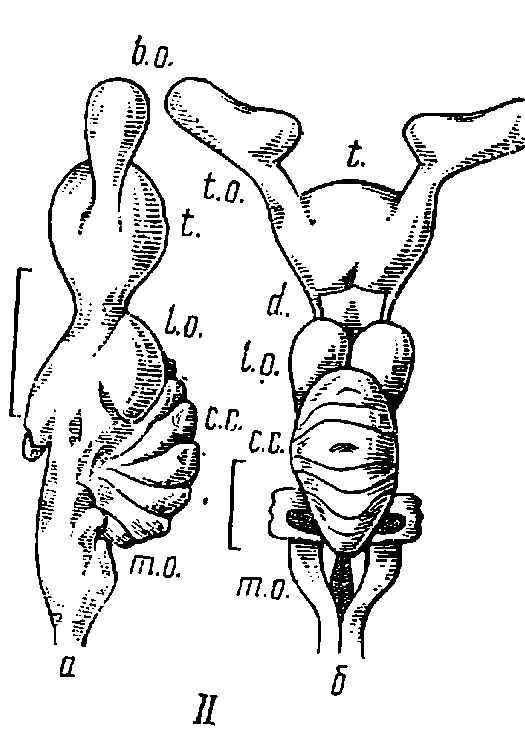
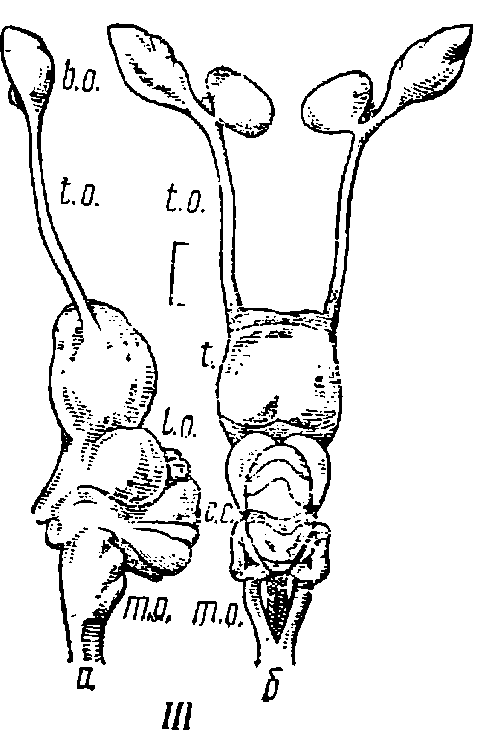
Рис. 6. Строение головного мозга акул.
I — кошачья акула (Scyliorhinus canicula) (мозг II типа); II — кунья акула (Mustelus canis) (мозг II типа); III — голубая акула (Prionace glauca) (мозг I типа); а — вид сбоку; б — вид сверху; шкала равна 10 мм; b. о. — обонятельные луковицы; t. о. — обонятельные тракты; t. — конечный мозг; d. — промежуточный мозг; l. о. — зрительные доли; с. с. — тело мозжечка; m. о. — продолговатый мозг.
За мозжечком расположен продолговатый мозг, который особенно хорошо развит у глубоководных акул, отличающихся высоким уровнем развития боковой линии [25, 73]. Продолговатый мозг современных акул (и скатов) плавно переходит в спинной мозг, представленный длинным округлым тяжем, проходящим под всем позвоночником акулы.
На нижней стороне головного мозга акулы расположены хорошо развитые нижние доли, сосудистый мешок и гипофиз. Эпифиз у акул слабо развит и лежит над промежуточным мозгом и каудальной частью переднего мозга.
От головного мозга акул отходит 11 пар черепных нервов, что отличает их от костистых рыб, имеющих только 10 пар черепных нервов. Черепные нервы соединяют мозг акул со всеми внутренними органами и органами чувств.
Развитие и соотношение отделов головного мозга акул тесно связано с особенностями их экологии (прежде всего питанием и местообитанием, причем данная морфоэкологическая связь хорошо прослеживается как на межвидовом, так и на внутривидовом уровнях.
Органы чувств. Акулы имеют хорошо развитые органы чувств. Бытовавшее до конца 60-х годов мнение, что у акул хорошо развито только обоняние и они являются своеобразными «плавающими носами», в корне опровергнуто последними исследованиями.
Обоняние у акул действительно очень хорошо развито, но ведущую роль оно играет лишь у акул-бентофагов (ряда видов семейства ScyLiiorhinidae и акул семейства Triakidae), полярной акулы (Somniosus microcephalia) и акул-молотов (Sphyrnidae). Органы обоняния у акул представлены парными ноздрями, расположенными на вентральной стороне головы (кроме плащеносной акулы и акул-молотов) перед ртом. Внутри ноздрей расположены обонятельные мешки, выстланные обонятельным эпителием. Снаружи ноздри прикрыты носовым клапаном различной формы, причем сложность формы и развития последнего говорит о большой роли обоняния в поведении акул, что зависит от экологии и прежде всего питания акул.
Помимо обоняния у акул хорошо развито зрение, особенно у видов, обитающих в толще воды. Зрение выполняет вспомогательную роль на расстоянии до 10—15 м (для различения силуэтов и движения объектов) и основную — на расстоянии до 3 м. Развитие зрения и глаз у акул также тесно связано с их экологией. Глаза акул имеют эллипсовидную форму с округлым хрусталиком и уплощенной роговицей. У ряда видов акул имеется мигательная перепонка. В отличие от скатов среди акул отсутствуют слепые виды.
Большое развитие у акул (особенно древних и ведущих глубоководный или придонный образ жизни) получила система органов чувств боковой линии. Она представлена у акул двумя типами образований: собственно боковой линией — каналом, проходящим по бокам вдоль всего тела акул, в стенках которого лежат скопления чувствительных клеток (у всех акул, кроме плащеносной, канал покрыт кожей и представлен внешне рядом отверстий); ампулами Лоренцини, расположенными на голове акул. Последние представляют собой небольшие капсулы с чувствительными волосками, погруженными в кожу и мышцы акул. Имеются данные (предварительные), что эти ампулы улавливают не только колебания воды, как собственно боковая линия, но и изменения ее плотности, солености и температуры.
Сравнительно хорошо развиты у акул органы слуха и вкуса, но их изучение еще только начинается. Однако уже сейчас можно говорить о связи развития этих органов чувств с их экологией.
Рекомендуем ознакомится: http://biofile.ru